植生班(徳島生物学会)
鎌田磨人1)・友成 孟宏2)・井内久利3)・
西浦宏明4)・石井愃義5)・森本康滋
1.はじめに
那賀川町は徳島市の南東約15km
の臨海部沖積平野上に位置しており、その南側には1級河川である那賀川が流れている(図1)。当地は、その面積1917ha
のうち、43%にあたる832ha
を耕地(水田が820ha,畑地が11ha,果樹園が1ha)が占める農業地域である(徳島県,1992)。全国的には、このような沖積平野の都市部に近接する地域は、宅地開発などにより景観構造が急変している傾向がある。那賀川町でも、国道55号線のバイパスの開通やゴルフ場建設などに伴い、急激な変化が起こると思われる。今後、農耕地環境、居住地環境、生物環境が有機的に結びつくように景観配置をし、生物群集を安定して維持できるような都市計画が望まれる。
近年、生態学的に安定した景観配置を行うための理論的な枠組みとして、景観生態学が発展してきている(たとえば
Forman and Godron,1986;Naveh and
Lieberman,1984;鎌田,1993)。また、アメニティ活動の一端として、生物群集の多様性を維持できるような農耕地や水辺環境を積極的に復元し、維持して行くような環境設計がスイスやドイツなどを中心に実践されており、日本でもこうした町づくりが期待されている(福留,1989;埼玉県野鳥の会,1990)。我々はこれらの問題意識に基づき、今後の那賀川町の調和的な町作りに役立つような景観生態学的研究としての基礎資料を得るために、町内の代表的な景観単位を抽出し、植生調査を行った。 この調査を行うにあたって那賀川町役場の方々から様々な便宜を図っていただいた。また、徳島県立博物館の太田陽子氏には調査資料のとりまとめをお手伝いいただいた。記して感謝する。
2.調査地の概況
1)自然環境
徳島県農林水産部(1979)によると、那賀川町の表層地質は、那賀川による三角州の未固結堆積物である砂礫(れき)、および粘土から成っている。土壌は、比較的内陸部には強粘〜粘質の細粒灰色低地土壌が、中間部には壌質土壌である灰色低地土壌が、臨海部には砂質あるいは礫質の粗粒灰色低地土壌や、排水不良土壌である細粒グライ、粗粒グライ、グライ土壌などが分布している。
気象環境については、那賀川町に関する資料が入手できなかったが、那賀川町に近い徳島市の暖かさの指数(WI)は131.8であり(鎌田ら,1991)、当地も気候区としては徳島市と同様に、常緑広葉樹林が成立する地域に含まれるものと考えられる。しかし、当地の土壌環境から、潜在自然植生は主にムクノキ−エノキ群集であるとされている(宮脇,1982)。
2)社会的環境
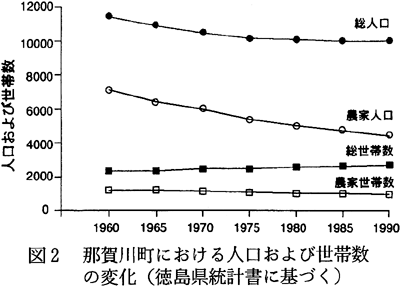
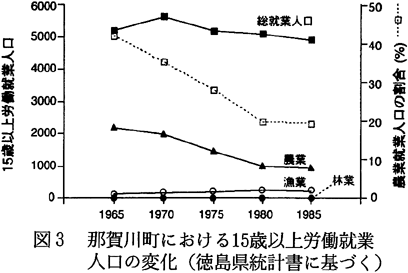
那賀川町は徳島市の近郊に位置しているために、ここ数年、人口では多少の減少が見られるものの、世帯数は増加しており、全体的には大きな変化は見られない。1960年の人口は11441人(2347世帯)であり、1990年は10006人(2727世帯)であった。しかしながら、その就業形態は変化してきている。すなわち、1960年に農家人口は7118人(1235世帯)と人口の62%を占めていたが、1990年には4469人(977世帯)と、全人口の45%にまで減少していた(図2.徳島県,1963〜1992)。15歳以上の労働就業人口では、その傾向がさらに顕著で、農業に従事する者は1965年には2193人(15歳以上就業人口の42%)であったのが、1985年には951人(19%)にまで減少していた(図3.徳島県,1963〜1992)。これは、農業から、二次・三次産業への職業の転換が起こったことによっている。
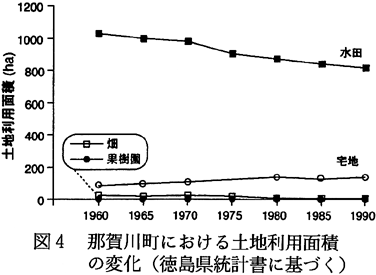
このような労働形態の変化につれて、土地利用形態も変化してきており、水田面積が減少する一方、宅地面積が増加してきている(図4.徳島県,1963〜1992)。また那賀川町内では国道55号線のバイパスが開通し、さらに臨海部にはゴルフ場が造成されつつある。こうした社会的な変化は、今後の土地利用のあり方を根本的に変化させるものであろう。すなわち、耕作地、水路などからなる田園的景観から、宅地などによって土地が優占される‘都市的’景観への変化が加速的に進行することが予想される。
3.調査の方法
調査の対象とした植生は以下のようなものであった。すなわち、那賀川河口の海浜砂丘植生、那賀川の河原植生、海岸沿いのクロマツ群落、および神社をとりまいている植生である。これらの植生の中で、Braun-Blanquet(1964)の方法を参考にして、草本群落については2m×2m、低木群落については5m×5m、高木群落については15m×15m
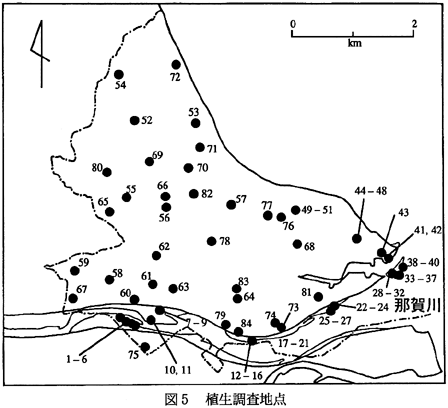
の方形区を基本として、その中に出現した維管束植物の被度および群度を階層別に記録した。なお、地形的な制約などから、方形区の面積はこの基本をはずれるものもあった。全調査地点数は84個所であるが(図5)、調査した神社の中には植被を持たないものが12個所あり、これらの地点は、解析の際には除外した。
現地調査は、1994年7月29〜31日、および8月3〜5日に行い、植物社会学的調査を行うとともに、植生の種類およびその広がりを1/25,000
の地形図に書き込んだ。そして、これらの調査結果をもとに1/25,000
の相観植生図を作成した。
植被を持つ72個の調査区で、出現した種を用いて群落の識別を行い、総合常在度表を作成した。さらに、識別された群落の類似性や独立性を検討するために、各調査区で出現した種およびその被度を用いて森下の類似度指数
Cλ(p)(Morisita,1959)を求めた。そして、さらに群平均法を用いてその類似度指数のクラスター分析を行った。
4.結果と考察
1)那賀川町の植物群落
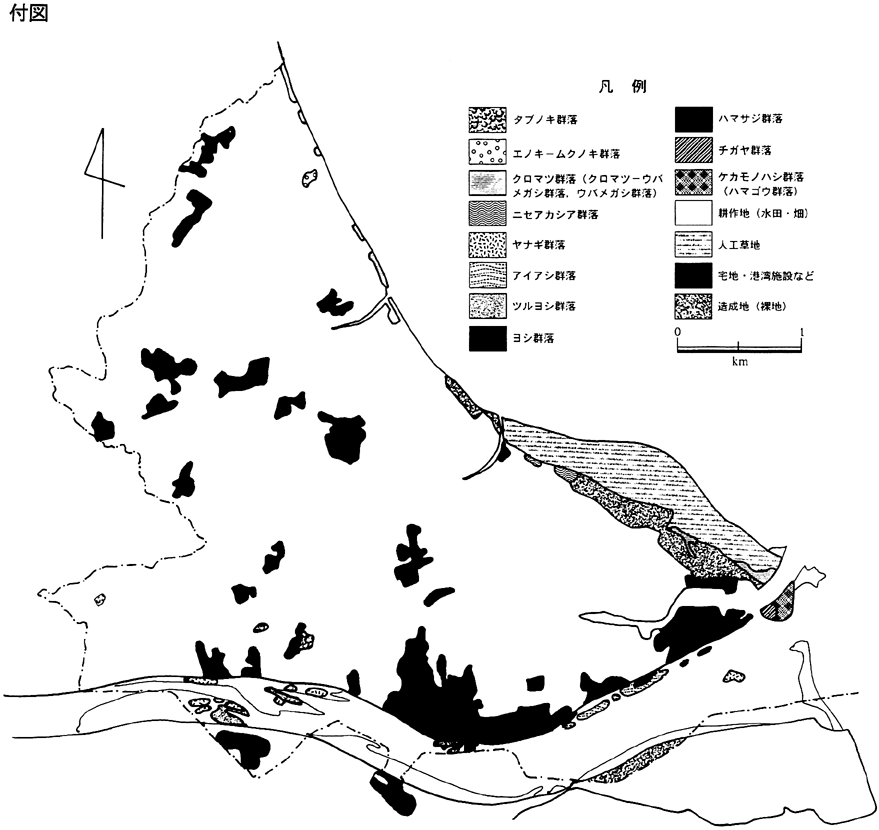
植被を持つ72個の調査区に出現した種からは、15の群落が区分された(表1)。すなわち、ケカモノハシ群落(省略記号;Ia)、ハマゴウ群落(Vr)、チガヤ群落(Ic)、シオクグ群落(Cs)、ヨシ群落(Pa)、ハマサジ群落(Lt)、ツルヨシ群落(Pj)、ヤナギ群落(Sa)、アイアシ群落(Pl)、ニセアカシア群落(Rp)、クロマツ群落(Pt)、クロマツ−ウバメガシ群落(PQ)、ウバメガシ群落(Qp)、タブノキ群落(Pet)、エノキ−ムクノキ群落(CA)であった。これら群落区分に基づいて描いた相観植生図を付図に示す。なお、クロマツ−ウバメガシ群落、ウバメガシ群落、およびハマゴウ群落は、その面積が小さかったために、植生図では表示できなかった。そのため前2群落はクロマツ群落に、ハマゴウ群落はケカモノハシ群落に含めて示した。その他、面積が小さいため調査、図示の対象から除いた群落もある。以下、区分された群落について若干の説明を加えておく。
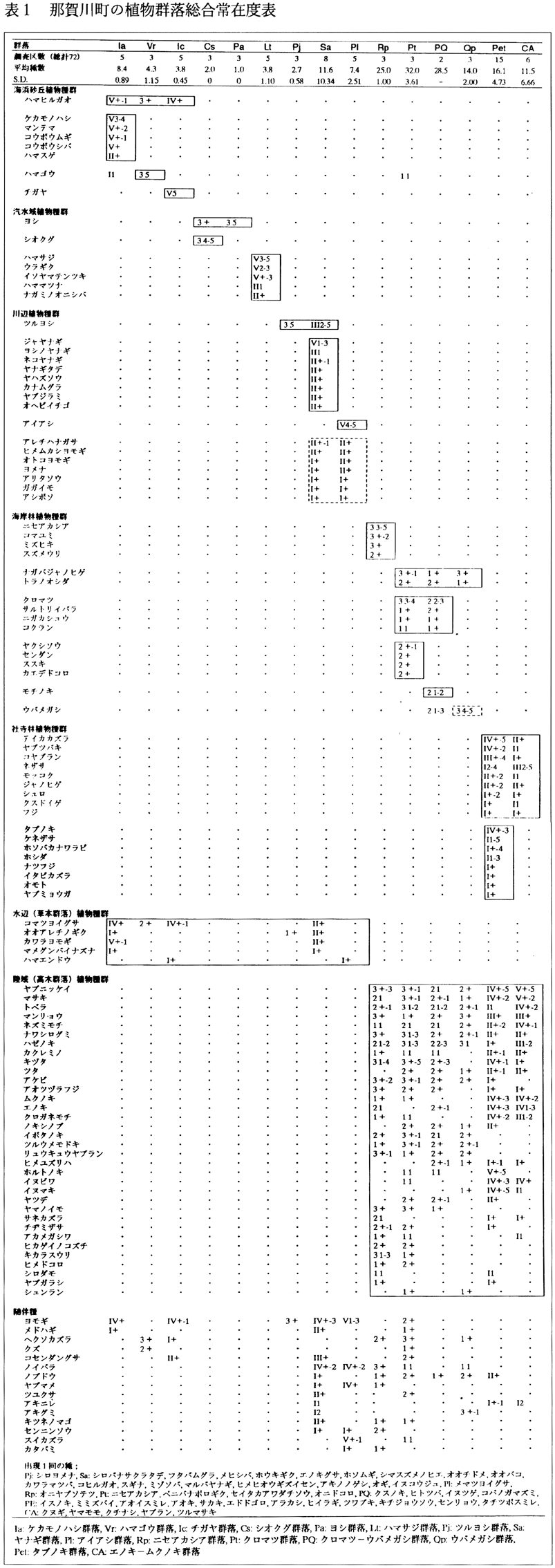
(1)ケカモノハシ群落
ケカモノハシ、マンテマ、コウボウムギ、コウボウシバ、ハマスゲで区分される。ケカモノハシが植被率25〜75%で優占しており、平均出現種数は8.4種であった。那賀川河口の造成地の砂丘上に成立していた。
(2)ハマゴウ群落
ハマゴウのみで区分される。ハマゴウの植被率は75%以上であり、平均出現種数は4.3種であった。那賀川河口のケカモノハシ群落に接して分布していた。
(3)チガヤ群落
チガヤのみで区分される。チガヤの植被率はほぼ100%であり、平均出現種数は3.8種であった。
ケカモノハシ群落、ハマゴウ群落、チガヤ群落では、ケカモノハシ群落が最も汀(てい)線に近い所に、チガヤ群落がその上部に、そして、ハマゴウ群落が最も高い所に分布していた。また、これら3群落にはハマヒルガオが出現し、海浜砂丘植物群落を特徴づけていた。
(4)シオクグ群落
シオクグのみで区分され、ほかにはヨシが生育していた。平均出現種数は2種であった。面積はごく小さく、那賀川河口域の堤防沿いの、満潮時には海水がかぶるような所に帯状に分布していた。本群落の立地は、海水の影響よりもむしろ半冠水の影響を強く受け、また土壌は、排水の悪い粘土を含んだ粘性の強い砂土である(宮脇,1982)。
(5)ヨシ群落
ヨシのみで区分される。ヨシの植被率は高く、ほぼ100%であり、他種は生育していなかった。那賀川河口域の砂質の州の上に分布していた。
(6)ハマサジ群落(図6)
ハマサジ、ウラギク、イソヤマテンツキ、ハママツナ、ナガミノオニシバで区分される塩沼植生。平均種数は3.8種で、那賀川のJR鉄橋下の州の上に分布していた。この群落を区分する種は、基本的に満潮時には塩水をかぶるような汽水域に生育するものである。ハマサジ群落生育地の土壌は砂礫を含むことが多く、礫間には砂質土壌がみられ、表層が粘土で覆われていることが多い(宮脇,1982)。ハマサジ群落は、かつて出島海岸にも見られたが、ゴルフ場造成により絶滅した。徳島県内の他の場所でも、河川改修・塩沼地埋立等の生育環境破壊・悪化に伴い、その分布地が極度に少なくなってきており、那賀川町内のこの群落は、分布上も、規模からも、徳島県内で非常に貴重な群落である。

(7)ツルヨシ群落
ツルヨシが75%以上の植被率で優占することによって区分される。平均出現種数は2.7種であった。那賀川JR鉄橋より上流の礫質の州の上に成立していた。
(8)ヤナギ群落
樹高が5m
以下のジャヤナギ、ヨシノヤナギ、ネコヤナギなどのヤナギ属木本や、ヤナギタデ、ヤハズソウ、カナムグラ、ヤブジラミ、オヘビイチゴによって区分される低木群落。林床にツルヨシが植被率90%以上で繁茂する群落も含まれる。平均出現種は11.6種であった。
(9)アイアシ群落
アイアシのみで区分される。平均出現種数は7.4種であった。那賀川河口域の砂質の州の上、平時は塩水をかぶらないような高位な所に成立していた。この群落が成立している州の、より低位な所にはヨシ群落が成立していた。
ヤナギ群落およびアイアシ群落にはアレチハナガサ、ヒメムカシヨモギ、オトコヨモギ、ヨメナ、アリタソウ、ガガイモ、アシボソなど路傍雑草・帰化植物が生育しており、河川植生の中では比較的乾燥した条件の中で成立していることがうかがわれる。
(10)ニセアカシア群落
ニセアカシア、コマユミ、ミズヒキ、スズメウリで区分される。優占種であるニセアカシアの樹高は20m
にまで達し、平均出現種数は25種であった。本群落はクロマツ群落の伐採後に植林して成立したものである。
(11)クロマツ群落
ヤクシソウ、センダン、ススキ、カエデドコロで区分される。また、本群落にはウバメガシが存在していない。調査したクロマツ群落には、クロマツの樹高が13m
程度にまで成長した群落と、6m
程度にまでしか成長していない群落が含まれる。平均出現種数は32種であった。
(12)クロマツ−ウバメガシ群落
モチノキのみで区分され、12m
程度にまで成長したクロマツの林冠下に、樹高6〜8m
程度のウバメガシが生育していることで特徴づけられる。平均出現種数は28.5種であった。 クロマツ群落およびクロマツ−ウバメガシ群落には、クロマツ、サルトリイバラ、ニガカシュウ、コクランなどが共通して出現する。
(13)ウバメガシ群落
明瞭な区分種は持たないが、ウバメガシが優占することで特徴づけられる。ウバメガシの樹高は7m
程度であった。群落内にはクロマツの切り株が残っており、クロマツ−ウバメガシ群落の林冠を形成していたクロマツが取り除かれたために成立した群落であると思われる。平均出現種数は14種であった。
本群落、クロマツ−ウバメガシ群落、クロマツ群落には、ナガバジャノヒゲ、トラノオシダが共通して出現した。
(14)タブノキ群落(図7)
タブノキ、ケネザサ、ホソバカナワラビ、ホシダ、ナツフジ、イタビカズラ、オモト、ヤブミョウガで区分され、群落高は10〜20m
であった。ホルトノキも多く、所によってはホルトノキが優占していた。一般にタブノキ林は、暖温帯の海岸部に成立する群落で、シイ林とともに極相林となりうる。しかし那賀川町内では、神社周辺のごく限られた小面積に成立しているに過ぎず、平均出現種数も16.1種と少ない。

(15)エノキ−ムクノキ群落
テイカカズラ、ヤブツバキ、コヤブラン、ネザサ、モッコク、ジャノヒゲ、シュロ、クスドイゲ、フジなどのタブノキ群落と共通する種を持つが、タブノキ群落を区分する種を欠くことによってのみ区分される。群落高は10〜15m
で、高木層にエノキやムクノキが優占する。平均出現種数は11.5種と、高木群落の中では最も少なかった。林内にシュロ、アオキ、ナンテン、コクサギなどを持つエノキ−ムクノキ群落は、地下水位が低いところに成立し、タブノキ林に近い立地的特徴を備えているという(宮脇,1982)。那賀川町で見られた本群落も、同様の立地的特徴を備えているものと思われる。
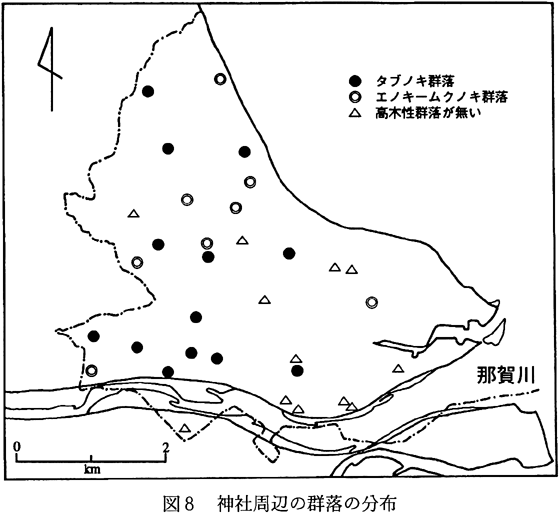
町内で調査した33個所の神社のうち、15個所の神社にタブノキ群落が、6個所の神社でエノキ−ムクノキ群落が分布していた。しかし、これらの多くでは、種組成や群落の構造などが貧弱であった。残りの12の神社では高木群落は存在していなかった(図8)。
2)群落間の関係
図9に森下の類似度指数
Cλ(p)に基づく、調査区間の類似度デンドログラムを示した。この図では、値が小さなものほど類似した構造を持っていることを示している。総合常在度表で区分された群落内は、比較的まとまったクラスターとして検出され、調査区の間で類似した構造を持つことが確認された。
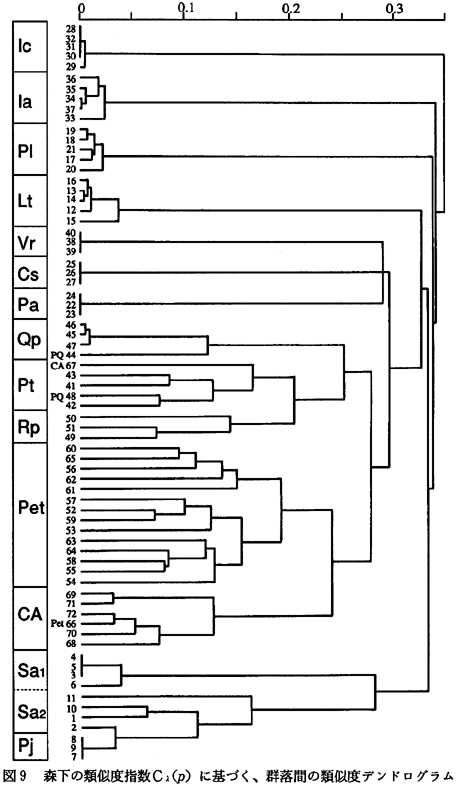
海浜砂丘植生であるチガヤ群落(Ic)、ケカモノハシ群落(Ia)、ハマゴウ群落(Vr)、汽水域植生であるヨシ群落(Pa)、シオクグ群落(Cs)、ハマサジ群落(Lt)、および川辺植生であるアイアシ群落(Pl)は、互いに他の群落との類似性が低く、独立した構造を持っていた。これは、これらの群落はハマゴウ群落を除いては草本群落であり、土壌条件や水分条件などの微細な環境の違いを反映して成立していることによると考えられる。換言すれば、海浜植生や塩沼植生などの多様性を維持するためには、それらの生育地の微細に異なる環境の多様性そのものが保たれる必要があることを示しているといえよう。
川辺植生であるヤナギ群落は、ツルヨシを欠く群落(Sa1)と、林床にツルヨシが優占する群落(Sa2)で構造的な違いがあるものの、ツルヨシ群落(Pj)とヤナギ群落については一つのクラスターを形成しており、比較的類似した構造を持っていた。
高木群落であるウバメガシ群落(Qp)、クロマツ群落(Pt)、ニセアカシア群落(Rp)、タブノキ群落(Pet)、エノキ−ムクノキ群落(CA)は一つのクラスターを形成していた。さらにその中では、ウバメガシ群落とクロマツ群落、および、タブノキ群落とエノキ−ムクノキ群落が比較的類似した群落として認められた。
ウバメガシ群落およびニセアカシア群落は、もともとはクロマツ林であった所に成立したものである。ウバメガシ群落やニセアカシア群落では、それらが成立する以前のクロマツ林の種構造を現在まで引き継いでいるために、構造的に類似した群落が成立したものと思われる。
クロマツ−ウバメガシ群落(PQ)は、ウバメガシ群落によって形成されるクラスターとクロマツ群落によって形成されるクラスターに分かれた。これは、クロマツ−ウバメガシ群落がクロマツ群落とウバメガシ群落との間の過渡的な群落であるためであろう。ただし、ここで区分されたクロマツ群落はウバメガシを欠く植分であった。原因は不明であるが、ウバメガシは海岸部から多少内陸によった、かつての自然堤防であると思われるような盛り上がった地形上のクロマツ林内に生育しており、海岸に隣接した所には存在していないようである。これは、海岸部の地形が成立した時期などの違いを反映しているのかもしれない。すなわち、自然落下、あるいはネズミなどの動物に種子の散布を頼るウバメガシは、より近年に成立した土地上には侵入できていないのかもしれない。実際、松茂町のように、1770年以降に埋め立てられてできた土地上に植林されたクロマツ群落内には、ウバメガシは侵入できていない(鎌田ら,1991)。
3)那賀川町におけるクロマツ林の変遷
現在、那賀川町において比較的まとまった高木群落を形成しているのは、クロマツ林である。那賀川町は、1956(昭和31)年に今津村と平島村が合併してできたが、以下に、合併以前の昭和初期および大正末期に刊行された「今津村史略(今津村,1930)」および「村史平島(平島村,1923)」から、このクロマツ林の変遷に関する事項を要約しておく。
「今津村史略(今津村,1930)」によると、藩政時代には江野島や今津浦にクロマツ林があったが、後者には番所もあり、ともに藩有林であった。江野島のクロマツ林は、1674(延宝2)年に手塚家三代助左衛門が植林を始めたことによってできたという。そして、それが1809(文化6)年には20ha(20町)余りのマツ林になっていたらしい。さらに、明治初期(1870年頃)には、マツの目通り直径が1m
程度(3〜4尺)にまでなっており、その林内には43戸の民家もあったという。
しかしながら、1875(明治8)年に開墾が行われたためか、1892(明治25)年の大暴風で砂浜や松林が壊滅的な打撃を受けた。このため、同年民家近くに石垣が築かれたが、その後も海岸の浸食が進んだため、1905(明治38)年から1916(大正5)年の間に、この一帯の海岸に堤防を築いた。これにより、堤防の外に残った43戸の民家は立ち退きを余儀なくされ、同時に、海岸にあったクロマツも放置され、徐々に強風や浸食などによって倒れていったらしい。この「今津村史略」を見る限りでは、明治末期から大正初期に築かれた堤防は、マツ林の内陸側あるいは林内に築かれたようである。堤防が築かれた当時には、堤防から波打ち際までの距離が300m(3町)もあり、畑地や砂浜があったという(今津村,1930)。
「村史平島(平島村,1923)」によると、1795(寛政7)年の苅屋村藪開検地帳、1862(文久2)年の上福井村林開下札、1830(文政13)年の工地村林開槓米麦帳などの存在から、かつてはそれ相応に樹林や竹林に富んでいたであろうと推測されている。藩政時代にはこれらの森林はすべて藩有となっていたが、ある一定の額を支払えば、名負林として出入りが許された。しかし、この場合でも上木の伐採は許されず、枝葉や下草、倒木などの採取が認められた程度であったという。これら採取された植物体は、燃料や肥料として利用されたのであろう。
1876(明治9)年における平島村内の林野面積は、原に0.06ha(6畝13歩)、中島に140.31ha(140町3反1畝22歩)、上福井に54.10ha(54町1反28歩)、工地に16.63ha(16町6反3畝15歩)、苅屋に2.86ha(2町8反6畝8歩)で、合計213.98ha(213町9反8畝26歩)であった。この林野には、いわゆる原野など、無立木地が含まれていると思われ、実際に森林としてみるべきものは、工地と上福井のマツ林のみであったという。工地のマツ林は、東西400m(4町)、南北520m(5町10間)程度、上福井のマツ林は東西600m(6町)、南北1200m(12町)程度あったという。どちらも民有林であり、樹高はそれほど高くなかったようである。
「村史平島」には、刊行当時の景勝地として、上福井の出島が挙げられており、「白砂青松」が映える「一幅画中」のものであるだけでなく、春には「ショウロ(松露)」、秋には「ハツタケ(初茸)」が採取できるキノコ山として、年中遊び客が絶えなかったと記されている。なお、ショウロは春に海辺砂地のマツ林内の地中に、ハツタケは秋にクロマツやアカマツの林内の地上に生えるキノコで、広く食用にされるものである。
これらのことから、昭和初期までの上福井のマツ林は、燃料や肥料として利用した枝葉や下草、そしてキノコなどの林産物資源を確保する場として、また、観光資源として重要であったことがうかがわれる。
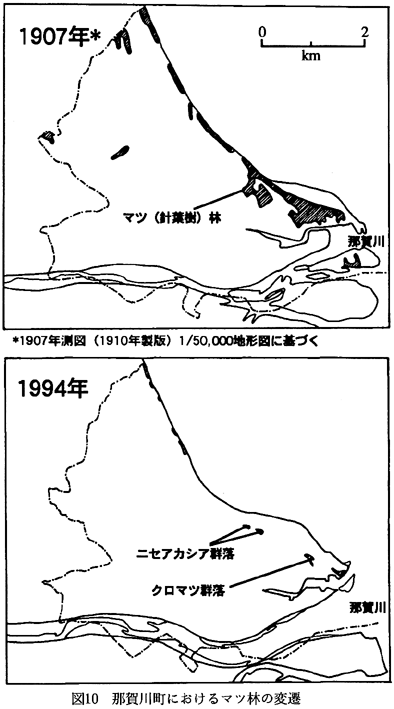
1907(明治40)年に測図された1/50,000
地形図から読みとったマツ林の分布と、今回の調査に基づいて作成した植生図から得たマツ林の分布を、図10に示した。なお、地形図に針葉樹記号で記されている場所はマツ林として読みとった。当地では他の針葉樹の分布は認められなかったからである。マツ林は、1907年には海岸沿いに連続して分布していたのに対して、現在ではごく一部に残存しているに過ぎない。これは、堤防の建設に伴ってマツ林が減少したことや(今津村,1930)、上福井出島付近では、ゴルフ場の造成や、宅地の造成によってマツ林が伐採されたことによると思われる。また工地の海岸部では、マツ林に代わってニセアカシア群落が成立している。
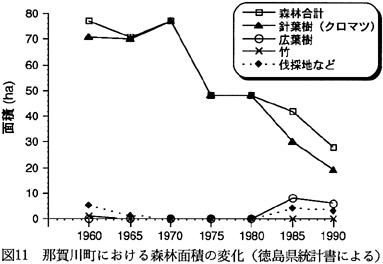
徳島県統計書(徳島県,1963〜1992)に基づいて、那賀川町における1960年以降の森林面積の変化を図11に示した。なお、徳島県統計書で針葉樹林、原野として区分されているものは、それぞれクロマツ林、および伐採地と考えてさしつかえなかろう。1960年以前の変化については今のところ不明であるが、当地でのクロマツ林の面積は、1970年より後に激減している。すなわち、1960年に71ha、1970年に77ha
であったマツ林の面積は、1990年には19ha(1960年の27%)にまで減少している。また、1980年より後にはマツ林に代わって、わずかではあるが、広葉樹林(ニセアカシア群落などであろう)や伐採地が増加しているのが特徴的である。
ここで見てきたように、那賀川町におけるクロマツ林は、防潮林として植栽され、林産物採取の場として、そして観光資源として、多目的に有効に利用されてきたものであった。しかしながら現在では、クロマツ林が果たしてきた機能は顧みられることもなく、放置されたり、伐採されている。
森林の少ない那賀川町においては、クロマツ林は景観の多様度を高め、生物群集の多様性を維持するために重要な空間である。また、臨海部の土地を守ってきた歴史を内在している森林であるともいえる。しかし、一般にマツ林は遷移初期段階の群落であり、その維持には積極的な管理が必要である。江野島、今津浦付近のクロマツ群落(図12)などは、そのクロマツ林をそのまま放置すると、林冠を形成しているクロマツが枯れるなどして取り除かれた場合には、群落の構造やそれが持つ機能は貧化すると思われる。このクロマツ群落内には、クロマツに代わって林冠を形成できる高木性の植物が生育していないからである。クロマツ林の持つ意味や維持の方法などについて、もう一度検討してみる必要があるのではなかろうか。

5.おわりに
以上、那賀川町における植生の分布、また、クロマツ林の変遷についてまとめた。すでに述べたように、那賀川町における土地利用形態は、農業的土地利用から都市的土地利用へと、劇的に変化して行くことが予想される。また、現在の那賀川町は、その植生の多様度や群落構造から見て、貧弱である。このような地域においては、現存の植生を保全しながら有効に利用して行くこと、さらに、積極的に植生の多様度を高めて行くことが重要であろう。その方法としては以下のようなことが考えられる。
まず、那賀川やその河口にある植生の保全である。特に、ハマサジ群落や、シオクグ群落などの塩沼植生は、その生育地が徳島県内では激減してきているものであり、何らかの保全の手段を講じなければならない。やむを得ず河川改修などの工事をする際にも、生育環境の保全に努めるべきである。また、ヨシ群落やツルヨシ群落、ヤナギ群落は、鳥類などの生息場所であることにも留意する必要がある。
次に、クロマツ林の持つ意味について再検討することである。クロマツ林は、防潮や防風の機能を持つだけでなく、多くの生物の生息空間でもある。さらに、それは防潮のために過去の人々が植林してきた歴史的な遺産であり、かつては、観光資源として、また林産資源の採取場所として機能してきたものであった。こうした意義からも、今後、防潮林や防風林としての機能を持った森林を整備・拡充する計画を策定し、現存するクロマツ林へのクロマツやウバメガシなどの補植にとどまらず、新たなクロマツ林の創造までをも行うべきであると考える。
最後に、内陸地で積極的な植栽を行うことであろう。神社周辺にはタブノキ群落やエノキ−ムクノキ群落が成立している所がある。これらの群落には当地の潜在自然植生を反映しているものもあると考えられるが、群落構造は貧弱なものが多い。また、植被のない神社も存在している。こうした神社の周辺を中心として、公園などに潜在自然植生を利用した植栽を行うことは、植生の多様度を高める上で有効である。
今後、今回取り上げた問題点が、様々な視点から検討され、さらに具体的な調査に基づいた、生態的に調和のとれた町づくりのためのプランが作られ、実施されることを期待する。
引用文献
Braun-Blanquet, J. 1964.
Pflanzensoziologie, Grundz■uge der Vegetations kunde. pringer,
Wien.
Forman, R. T. T. and Godron, M. 1986. Landscape ecology. Wiley,
New York.
福留脩文.1989.スイスの河川工事と生物保護.淡水魚保護,No.
2:49-52.
平島村.1923.村史平島.平島村(那賀川町,1985再発行).
今津村.1930.今津村史略,全.今津村役場.
鎌田磨人・森本康滋・石井愃義・友成孟宏・西浦宏明・井内久利.1991.松茂町の植生.総合学術調査報告 松茂町(郷土研究発表会紀要37号),23-37.阿波学会・徳島県立図 書館,徳島.
鎌田磨人.1993.「景観生態学」のめざすもの.国際景観生態学会日本支部会報,1:31- 40.
宮脇 昭編著.1982.日本植生誌 四国.至文堂,東京.
Morisita,
M. 1959. Measuring of interspecific association and similarity between
communities. Mem. Fac. Sci ., Kyushu Univ., Ser. E. (Biol.) ,
3:65-80.
Naveh, Z. and Lieberman, A. S. 1984. Landscape ecology, Theory
and application, Springer-Verlag, New
York.
埼玉県野鳥の会.1990.ビオトープ,緑の都市革命.ぎょうせい,東京.
徳島県.1963,1966,1972,1977,1982,1987,1992.徳島県統計書(昭和35年,昭和40 年,昭和45年,昭和50年,昭和55年,昭和60年,平成2年).徳島県,徳島.
徳島県農林水産部.1979.土地分類基本調査,阿波富岡.徳島県,徳島.
1)徳島県立博物館 2)阿南市立福井南小学校 3)徳島県立川島高等学校
4)徳島県立池田高等学校 5)徳島大学総合科学部