植生班
鎌田磨人1)・森本康滋2)・
石井愃義3)・友成孟宏4)・
西浦宏明5)・井内久利6)
1.はじめに
松茂町は徳島市に隣接し、臨海部の沖積平野上に位置する農業地帯で、江戸時代中期以降の新田開発により発達してきた地域である(阿子島 1975,岸本 1969;図1)。

全国的には、このような沖積平野の都市部に近接する地域は、宅地開発などにより、景観構造が急変している傾向がある。当地は、徳島市近郊に位置するにもかかわらず、‘田園的’景観をとどめている。また、海岸、水路など水辺環境に富む地域でもある。
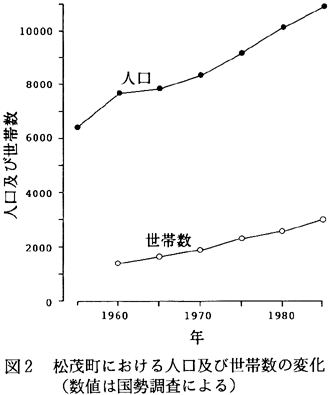
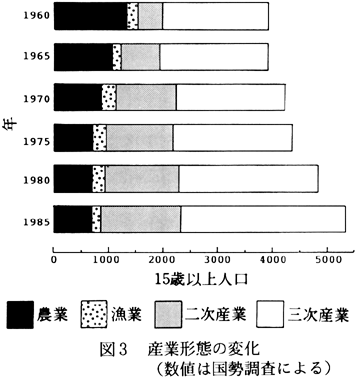
しかし、全域が平坦であること、徳島市に隣接していることなどから、近年は宅地開発などにより人口および世帯数は年々増加してきている(図2)。また、農業従事者の減少、二次・三次産業従事者の増加など、産業形態が変化してきている(図3)。このような人口、産業の変化につれて、土地利用形態も変化してきており、耕地面積が減少する一方、宅地面積が増加してきている(図4)。今後さらに、耕作地、水路などからなる田園的景観から、宅地などによって土地が優占される‘都市的’景観へと変化していくことが予想される。今後、農耕地環境、居住地環境、生物環境が互いに結び付き適合するように景観配置をし、生物群集を維持していくことが望ましい。

近年、この景観配置の理論枠として景観生態学が発展してきている(例えば
Forman & Godron 1986, Naveh & Lieberman 1984, Turner
1989)。またアメニティ活動の一端として、生物群集の多様性を維持できるような農耕地や水辺環境を積極的に創造し、維持していくような環境設計が、スイスやドイツなどを中心に実践されている(福留 1989,埼玉県野鳥の会 1990)。
我々は、今後の松茂町の調和的な町作りに役立つような景観生態学的研究としての基礎資料を得るために、町内の代表的な景観単位を抽出し、植生調査を行った。その結果、若干の知見を得たのでここに報告する。
2.調査地の概況
徳島県農林水産部(1987)によると、松茂町の表層地質は未固結堆積物の砂(海成層)であり、海岸部の一部に砂礫層が、今切川沿いの一部に粘土―シルトからなる地質が存在している。土壌は、海砂によって造成された粗粒灰色低地土壌や、排水不良地に分布する水田土壌である粗粒グライ土壌などから成る(徳島県農林水産部 1987)。
気象環境については、松茂町に関する資料が入手できなかったが、松茂に隣接する徳島市の資料を基にした、気候図形を図5に示した。

気候図型は、1年間の月平均気温と月雨量の変化曲線からなる。縦軸の目盛りを、気温の30℃が雨量の
60mm
に一致するようにきざむと、気温曲線より雨量曲線のほうが下に位置する時期が、自然の植物にとって決定的に水の不足する時期であることが経験的に知られている(Walter
1973)。したがって、雨量曲線と気温曲線の間の面積の大きさが、植物にとっての水ストレスの大きさを表すと考えられる。一方、平均雨量が月 100mm
以上ある季節は、まず水不足は起こらない。
さらに、徳島市の資料を基にして算出した、吉良の暖かさの指数(WI)および乾湿度指数(K)を、表1に示した。一般に、85<WI<180
であるような場所には、常緑広葉樹林が成立すると言われている(吉良 1976)。

これらのことから判断すると、松茂町は気候区としては非常に暖かく、常緑広葉樹林帯に属していると考えられる。しかしながら、乾湿度指数から見ると、その値(9.5)は小さく、特に、冬季の乾燥は植物の生育にとって不利であると思われる。
3.調査の方法
現地調査は、1990年8月2日から4日にわたって行った。
松茂町の代表的な植生および土地利用は、クロマツ林、海岸植生、果樹園(ナシ)、水田、ハス田、畑地(サツマイモなど)、造成地、宅地などであった。これらの景観単位の中から、図6に示される地点で植生調査を行った。図6に示された番号の内、1〜3が海岸植生、4〜8が海岸部クロマツ群落、14が内陸部クロマツ群落(並木)、9〜13が造成地、15がハス田、16がナシ果樹園、17が水田である。
海岸植生の調査に当たって、調査地点番号1では、汀線から内陸に向かう直線上で、1m
間隔で1m×1m
の方形区を計81個設け、その枠内に出現した維管束植物の種類と植被率を調べた。その他の調査区では、同様のトランセクトで種類のみを調べた。海岸部クロマツ群落では、10m×10m
の方形区内に、造成地の草本群落では3m×3m
の方形区内に出現した全維管束植物の種類と植被率を調べた。これら4植生の調査は、Braun-Blanquet(1964)の植物社会学的方法に基づいて行った。
内陸部のクロマツ群落では、十分な幅で方形区を設けられなかったので、群落内に出現した植物の種類と階層を、長さ約
100m
にわたって調べた。また、ハス田、ナシ園、水田では、その耕作地内に入ることがはばかられたので見取り調査を行い、植物相を調べた。
これらの調査結果、1986年空中写真(国土地理院撮影)の判読および1988年の
1/25,000
地形図(国土地理院発行)の凡例などを参考に、相観による植生図を作成した。

4.結果と考察
(1)造成地の草本群落
造成地の草本群落についての種組成表を、表2(表右)に示した。
造成地の草本群落は、帰化植物であるセイタカアワダチソウ(Solidago
altissima)が優占するセイタカアワダチソウ群落としてまとめられた。造成地など、裸地化された場所に帰化植物が繁茂することは、一般的な傾向として指摘されている。松茂町は、現在造成中の場所が多く分布しており、そうした開発にともない本群落の分布場所およびその面積は増加してきていると考えられる。造成後にその土地が放置された場合は、おそらくはクロマツなどの陽生木本が侵入してくると思われるが、多くの場合は工場地や宅地として利用され、生態学的には退行遷移が起こる。工場地開発、宅地開発を行う場合には、その周辺に空き地などを配置し、潜在自然植生を念頭に置き、遷移系列を利用する方法、すなわち後記クロマツ群落の遷移に示されるような流れを利用して緑化を進め、緑地を確保することが望ましい。
松茂町においては、宅地、工場地、緑地の調和的な配置およびその方法が、重要な課題としてとりあげられるべきである。

(2)耕作地
耕作地であるハス田、水田、ナシ果樹園(図7)における植物相を表3に示した。

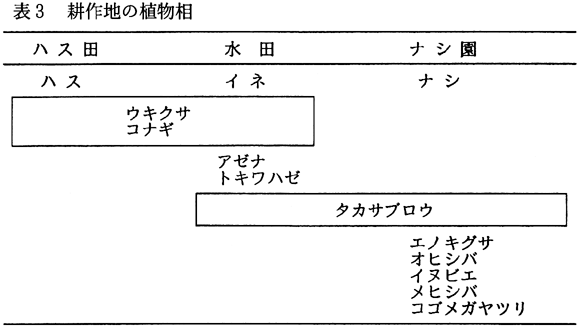
それぞれの耕作地で優占していたのは、作物であるハス(Nelumbo
nucifera)、イネ(Oryza sativa)、ナシ(Pyrus
serotina)であった。ハス田、水田では、その水環境を反映して、ウキクサ(Spirodela polyrhiza)、コナギ(Monochoria
vaginalis)が存在していた。水田では、畑地の雑草でもあるトキワハゼ(Mazus pumilus)、アゼナ(Lindernia
procumbens)が出現した。また、タカサブロウ(Eclipta
prostrata)が水田、ナシ園にわたって出現した。ナシ園では、畑地雑草のエノキグサ(Acalypha
australis)、イヌビエ(Echinochloa crus-galli)、メヒシバ(Digitaria
adscendens)、オヒシバ(Eleusine indica)、コゴメガヤツリ(Cyperus
iria)が存在していた。これらの耕作地に出現する植物の種数は極めて少なく、農薬散布、雑草取りなどの影響を反映していることがうかがわれた。
自然植生の少ない農村や都市部では、耕作地そのものが生物の多様性を維持していくための重要な空間となるが、作物収穫の経済性、管理のしやすさの面から、画一的な管理(例えば農薬散布)が行われやすい。しかしながら、耕作地が身近な生物たちの重要な生息地であり、また、その生物たちが住民に憩いを提供するという側面を見落とさないような配慮が必要であろう。そのような視点で、生物的防除(総合防除)に関する基礎的研究が行われており(桐谷・中筋 1977)、さらに近年、天敵を利用した生物的防除、無農薬農業の可能性が示唆されるに至っている(高橋 1989,日鷹 1989,日鷹・中筋 1990)。
果樹園も、生物、特に鳥類にとっての生息場所、猛禽類の待ち伏せ場所、さえずりの場所、また一般に外敵・悪天候からの避難所、越冬の場所などとして重要である。農薬、機械などを用いた集約的な管理が行われている果樹園と、粗放的な管理が行われている果物畑を比較した場合、粗放的な管理が行われている果物畑で、生物相が多様であることが指摘されている(Blab 1986)。また、その生息地としての果物畑の保全・保護の方法なども、ドイツなどでは実際に検討されている(Blab 1986)。
作物としての収量や品質を保つことは重要であるが、作物の生物学的被害(収量・品質)の許容範囲を正確に見極め、それに応じた管理を行うことが重要であろう。それには、生産者の努力だけでなく、消費者の考え方、品質の捉え方などをも変革させる必要がある。松茂町がそのような情報の発信源となることが期待される。
(3)海岸植生
調査した3地点で出現した全種は、以下の通りであり、総種数は27種であった。コウボウムギ、オカヒジキ、ハマヒルガオ、コマツヨイグサ、ケカモノハシ、ヒメムカシヨモギ、オオフタバムグラ、ハマボウフウ、オオアレチノギク、ヨモギ、チガヤ、ヘクソカズラ、セイタカアワダチソウ、ノイバラ、ススキ、メヒシバ、ツルナ、イヌホオズキ、ギシギシ、ハマエンドウ、ハマスゲ、キシュウスズメノヒエ、コウボウシバ、アツバキミガヨラン、アキノノゲシ、スゲ属の一種、クロマツ。

海岸における植物の分布の様子を、調査地点番号1の結果を基にまとめたのが図8である。横軸は汀線からの距離、縦軸はそれぞれの植物の各地点における植被率の階級を示してある。海岸部では、汀線からの距離に応じて、植物の種類に変化が生じていた。すなわち、コウボウムギ(Carex
kobomugi)は最も汀線に近い 30〜40m の地点で最も優占し、ハマヒルガオ(Calystegia
soldanella)、ケカモノハシ(Ischaemum anthephoroides)が 40〜70m に、コマツヨイグサ(Oenothera
laciniata)が 40〜100m に、オオフタバムグラ(Diodia teres)が 70〜100m に、最も内陸部の 90m
からはチガヤ(Imperata
cylindrica)などが優占していた。これは、それぞれの植物の塩分耐性の違いや、砂の移動量およびその変動環境への植物の適応の違いなどに由来すると思われる。
森本が1975年に同地点で調査を行った際には、コウボウムギ生育帯よりもさらに汀線側に、ギシギシが生育していた(森本 1978)。本調査では、ギシギシは内陸部の一部にみられただけで、1975年時点に一次的に定着していたものと思われる。
当地の海岸の砂浜部を、飛行場が拡張される以前と以降で比較すると、砂の堆積の様子が変化してきている。すなわち、1961年に国土地理院が撮影した空中写真および1978年発行の
1/25,000 地形図と、1986年の空中写真および1988年発行の 1/25,000
地形図を比較すると、飛行場の北側に砂の堆積が見られ、砂丘の面積が拡大している。松茂町付近では播磨灘側からの沿岸流(あるいは恒流)が流れており(殿谷 1979,小坂 1985)、この沿岸流によって運搬されてくる砂が、飛行場が張り出した地点でトラップされるようになったためと考えられる。今後、この砂の供給形態の変化につれて、植生配分も何らかの影響を受けることが予想される。
(4)クロマツ群落
海岸部クロマツ群落についての種組成表を、表2(表左)に示した。
このクロマツ群落は、クロマツ(Pinus
thunbergii)、マサキ(Euonymus japonicus)、トベラ(Pittosporum tobira)、イヌビワ(Ficus
erecta)などが優占する群落であった。宮脇(1982)によれば、四国の海岸部には、ハマビワ、ウバメガシを欠き、トベラ、マサキ、ハマヒサカキ、モチノキなどが優占する林分があり、トベラ、マサキ、ハマヒサカキ、オニヤブソテツ、クロマツ、シャリンバイなど海岸風衝地のヤブツバキクラス林を標徴あるいは区分する種群が生育していることにより、マサキ−トベラ群集としてまとめられるとしている。植林から出発した本群落は、この群集の要素を多く含むことから、植物社会学的には現在マサキ−トべラ群集に近くなっていると考えられる。
本群落は、エノキ(Celtis
sinensis var. japonica)、ハゼ(Rhus succedanea)、アカメガシワ(Mallotus
japonicus)などの陽生木本の種群が存在する林分と、それらを欠く林分に区分された。前者は、海岸沿いの北部(調査区番号6,7,8)に分布していた。
海岸部クロマツ林は、新田開発に伴って、防風、防潮などのために植栽されたものである。南部の豊岡地区では、1805年に一応の新田築立工事が完了し、豊岡新田と名付けられた。その際、1804年に松苗20万本が植えられたという(平島 1975)。一方、北部の満穂地区は、1820年に一応の工事が終了し、1884年に墾田が完成し、満穂新田と名付けられたという(平島 1975)。このように、北部と南部では新田完成時期に大きな開きがあり、クロマツの植栽時期も大きく異なっていたことが予想される。1961年国土地理院撮影の空中写真の判読から、当時は、北部の方でクロマツの樹高が低い傾向が認められた。前述の南部と北部における群落構造の違いは、このような群落成立の時期の違いに由来している可能性があると思われる。すなわち、成立年代の遅い北部の群落では、遷移初期に出現する陽生木本がまだ残っているものと思われる。倉内(1969)は、干拓地においてクロマツ林からタブ林、スダジイ林と遷移して行く中で、クロマツ林は成立から100年をピークに衰退していくことを示している。本群落は同じ遷移系列ではないが、この結果から推察すると、クロマツ群落としては衰退期に入っていると思われる。
クロマツ群落は、高木層(I層)には植被率10〜20%程度で、亜高木層(II層)には60〜90%の植被率でクロマツ(Pinus
thunbergii)が生育していたが、低木層(III層)及び草本層(IV層)にはクロマツは見られなかった。低木層には、マサキ(Euonymus
japonicus)やトベラ(Pittosporum
tobira)などが生育しており、その植被率は南部では80〜90%、北部では40〜50%であった。すなわち、南部の群落の方がマサキ−トべラ群集(宮脇 1982)により近くなって安定していることが示唆される。
海岸植生から海岸部クロマツ群落につながる景観の模式断面図を、図9に示した。海岸部の景観は、コウボウムギ(Carex
kobomugi)、ハマヒルガオ(Calystegia soldanella)、コマツヨイグサ(Oenothera
laciniata)、オオフタバムグラ(Diodia teres)、チガヤ(Imperata
cylindrica)、セイタカアワダチソウ(Solidago
altissima)などが連続的に出現する海岸から、道路をはさんでクロマツ林につながる景観単位として位置づけられた(図10)。


しかしながら、前述したように、海岸部クロマツ林の林床にはマサキやトベラなどの常緑性の低木が繁茂しており、陽生木本であるクロマツの稚樹は生育していなかった。また、当地のクロマツ群落は衰退傾向にあると考えられた。したがって、このまま放置すればクロマツは更新することなく消滅し、マサキ、トベラなどの低木群落に推移し、海岸部の景観も変貌する可能性があると思われる。
海岸部のクロマツ群落は防風などの防災的な役割のほか、日本人が海岸の美的な捉え方をする際の「白砂青松」としての海岸景観の重要な要素として、さらに当地では、新田開発の歴史などを内在する文化的景観として、大きな意味を持つと思われるが、それを保存するためには何らかの対策が必要となろう。
松茂町には、海岸部だけでなく内陸部にもクロマツ群落が存在し、クロマツ並木を形成していた(図11)。内陸部のクロマツ群落の分布は、新田を閉め切る堤防の位置(阿子島 1975)によく一致し、その堤防沿いに植林されたものと思われる。
内陸部クロマツ群落およびその周辺の模式断面図を、図12に示した。本群落を含む景観単位は、水路、盛土上のクロマツ群落、道路、ハス田などからなり、いわゆる田園的景観を特徴付ける景観単位として位置づけられよう。

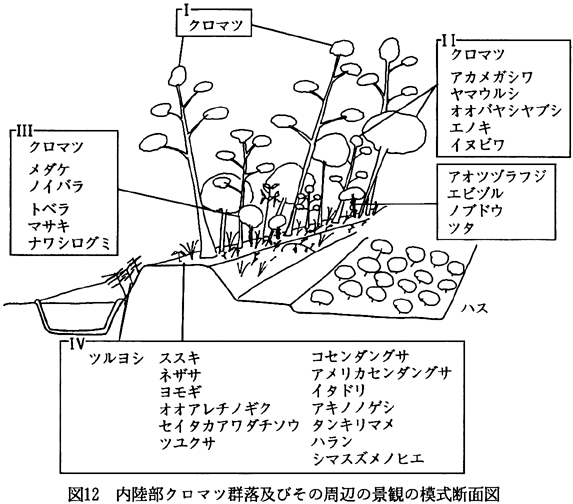
クロマツ群落は、高木層(I層)、亜高木層(II層)、低木層(III層)それぞれにクロマツ(Pinus
thunbergii)が生育していた。また、亜高木層には、アカメガシワ(Mallotus japonicus)、ヤマウルシ(Rhus
trichocarpa)、オオバヤシャブシ(Alnus sieboldiana)、エノキ(Celtis sinensis var.
japonica)、イヌビワ(Ficus erecta)などの陽生木本が、低木層には、メダケ(Pleioblastus
simonii)、ノイバラ(Rosa multiflora)などの陽生種、およびマサキ(Euonymus
japonicus)、トベラ(Pittosporum tobira)、ナワシログミ(Elaeagnus
pungens)などが生育していた。さらに、草本層には、ツルヨシ(Phragmites japonica)、ススキ(Miscanthus
sinensis)、ネザサ(Pleioblastus chino var. viridis)、ヨモギ(Artemisia
princeps)などの陽生草本類が生育していた。本群落は、低木層以下にもクロマツが生育していること、亜高木層、低木層および草本層に陽生種が多く生育していること、群落内に出現する種数が多いことなどで、海岸部クロマツ群落と異なっていた。これは、クロマツ群落が成立している盛土の幅が約2mと狭いために、良好な光環境が維持されていること、耕作地に隣接し人手が入りやすいことなどによっていると思われる。このような群落では、適当な頻度で下草刈りを行うことでクロマツを更新させることができ、その群落を維持し得ると考えられる。
海岸部クロマツ群落および内陸部クロマツ群落は、新田開発にともなって植林され、成立したものであった。植物は自分自身では移動できないので、クロマツ群落内に生育する植物は、何らかの要因でその種子が運搬され移入してきたものと考えられる。表4にクロマツ群落内(海岸部および内陸部)に存在していた木本の種子散布形態を示した。クロマツ以外のほとんどの樹種は、その種子が動物によって分散する型のものであった。特に、鳥によって分散する種が多かった(中越 1988,中西 1990,由井 1988)。このことは、植林によって成立したマツ並木であっても、その空間を利用するのは人間だけでなく、止まり場所、営巣場所、採餌場所などとして鳥類もまた利用している空間であることを示していると考えられる。マツ並木は新田開発の歴史を内在した空間であるというだけでなく、生物群集の多様性を維持していくための重要な空間であるといえよう。

しかしながら、1966年の空中写真から判読したクロマツ群落(並木)から比較すると、クロマツ群落は減少している。特に、宅地開発が進んでいる地区では、盛土が消失し、クロマツの密度も減少しているように思われる(図13)。クロマツ並木の積極的な保全は、松茂町の発展の歴史を地表面に残すことを意味する。そうした歴史性を町の景観作りに利用し、さらに新たな緑地空間としての並木を配置していくことは、今後、住民と生物が調和的に生きていくためにも必要なことではないだろうか。積極的な取り組みが期待される。

(5)相観植生
松茂町の現存植生について、相観によって区分した植生図(1/25,000)を付図に示した。この相観植生図では、水田とハス田を区分することが困難であったので、一括して示した。本植生図からも明らかなように、松茂町は水田・ハス田、畑地などの耕作地が卓越しており、そうした耕作地の中に新しい宅地空間が形成されつつある。セイタカアワダチソウ群落となっている造成地も、広い範囲を占めている。樹林地はクロマツ群落および果樹園以外になく、その面積も非常にわずかである。
このような植生配分の中で、前述してきたような緑地の保全を計り、生物群集の多様性を高めるような配慮が、今後の町づくりの過程で必要であろう。
5.おわりに
本報告では、耕作地、造成地、海岸植生、海岸部クロマツ群落、内陸部クロマツ群落を中心に、松茂町の植生を概観するとともに、今後配慮されるべき問題点について述べた。
本論で取り上げられなかった松茂町のもう一つの特徴は、豊富な水辺環境である。農耕地の周囲には多数の水路が設けられている。しかしながら、こうした水路はコンクリートで固められていて、生物にとっては利用しにくい空間になっている。管理の方法など検討すべき点は多いものの、例えば石積みの護岸の建設の方法などが考慮されるべきであろう。石積みによる護岸により、生物の分布密度、種数が劇的に増加することが指摘されている(Blab 1986)。また、河川幅などによっても規定されるが、ツルヨシなどの植物による、水鳥のための生息場所設計、また有機物に対する浄化効果なども考慮されてよいのではなかろうか。耕作地、宅地、工場地が混在する松茂町では、農薬、家庭排水、工場排水などで河川環境の悪化が心配される。特に、家庭排水による小水路の生物相への影響は継続的にチェックしていく必要があろう。
耕作地や宅地の周辺に、歴史的な特性を活かした並木を配置していくこと、その際、内陸部クロマツ群落(並木)でみられたように、他の樹種の侵入を許すような2m程度の盛土あるいは表土を残すことなどが、生物の多様性を高める上で重要であろう。また、広範囲にわたる造成地、特に工場地の周辺あるいは内部に空き地を残し、遷移にまかすことにより、緑地を創造していくような方法も模索されて良い。短期間での緑化にはならないが、長期的な視点に立った場合は、その土地に最も適した樹種による緑化方法となり、公園や野外教育の場として役立てられよう。
本報告書は、極めて短期間の調査に基づいて作成された。今後、今回取り上げた問題点が、様々な視点から検討され、さらに具体的な調査が成されるべきであろう。なにごとも経済優先の社会ではあるが、生態学的に調和のとれた町づくりが、この松茂町で模索されることが期待される。
引用文献
阿子島 功.1975.松茂の自然,地形・地質.「松茂町誌,上巻」(徳島県板野郡松茂町誌編纂委員 会編),36-53.松茂町誌編纂室,徳島.
Blab,
J. 1986.Grundlagen des Biotopschutzes f■r Tiere. Schriftenreihe f■r
Landschaftspflege und Naturschutz, Nr.
24.(武内和彦・大黒俊哉訳編.1989.ビオトープと動物保護.)
Braun-Blanquet, J.
1964.Pflanzensoziologie, Grundz■ge der Vegetationskunde. 3. Aufl.
Springer-Verlag, Wien and New York.
Forman, R. T. T & Godron, M.
1986.Landscape ecology. Wiley, New
York.
福留脩文.1989.スイスの河川工事と生物保護.淡水魚保護,no.2:49-52.
日鷹一雅.1989.自然農法稲作と集約的農法稲作における生物群集の構造および機能の比較研究,学位(博士)論文要旨.広島大学総合科学部紀要VI,no.15:85-88.
日鷹一雅・中筋房夫.1990.自然・有機農法と害虫.冬樹社,東京.
平島顕吉.1975.地区の歴史.「松茂町誌,上巻」(徳島県板野郡松茂町誌編纂委員会編),3-30.松茂町誌編纂室,徳島.
吉良竜夫.1976.陸上生態系―概論―,生態学講座2.共立出版,東京.
桐谷圭二・中筋房夫.1977.害虫とたたかう―防除から管理へ.日本放送出版協会,東京.岸本 實.1969.徳島県総説,II歴史的背景.「日本地誌18,香川県・愛媛県・徳島県・高知県」(青野壽郎・尾留川正平編),302-305.二宮書店,東京.
小坂淳夫編.1985.瀬戸内海の環境.恒星社厚生閣,東京.
倉内一二.1969.植物群落の遷移.「図説植物生態学」(沼田 真編),129-188.朝倉書店,東京.
宮脇 昭編著.1982.日本植生誌,四国.至文堂,東京.
森本康滋.1975.徳島県の海岸植生、徳島県立城南高等学校研究紀要,no.4:17-34.
中越信和.1988.種子集団の生態2,散布のしくみとその過程.日本の生物,2(2):46-49.
中西弘樹.1990.海流の贈り物,漂着物の生態学.平凡社,東京.
Naveh,
Z. & Lieberman, A. S. 1984.Landscape ecology, Theory and application.
Springer-Verlag,New
York.
埼玉県野鳥の会.1990.ビオトープ,緑の都市革命.ぎょうせい,東京.
高橋史樹.1989.対立的防除から調和的防除へ.農村漁村文化協会,東京.
徳島県農林水産部.1987.土地分類基本調査,徳島.徳島県.
殿谷次郎.1979.大型冷水塊形成による黒潮流及び徳島沿岸の海況変動.徳島県水産試験場事業報告,128-135.
Turner,
M. G. 1989.Landscape ecology, the effect of pattern and process. Annu.
Rev. Ecol. Syst., 20:171-197.
Walter, H. 1973.(translated by Wieser, J)
Vegetation of the earth, in relation to climatic and the eco-physiological
conditions. Springer-Verlag, New York.
由井正敏.1988.森に棲む野鳥の生態学.創文,東京.

1)徳島県立博物館 2)阿南工業高等学校 3)徳島大学総合科学部 4)芝坂小学校 5)池田高等学校 6)川島高等学校