| 阿波学会研究紀要 |
| このページでは、阿波学会研究紀要論文をご覧いただけます。 なお、電子化にともない、原文の表記の一部を変更しています。 |
| 郷土研究発表会紀要第11号(総合学術調査報告 鳴門) | |
| 鳴門市における人体寄生虫とその中間宿主の分布について | 生物学班 豊岡磊造 |
|
鳴門市における人体寄生虫とその中間宿主の分布について
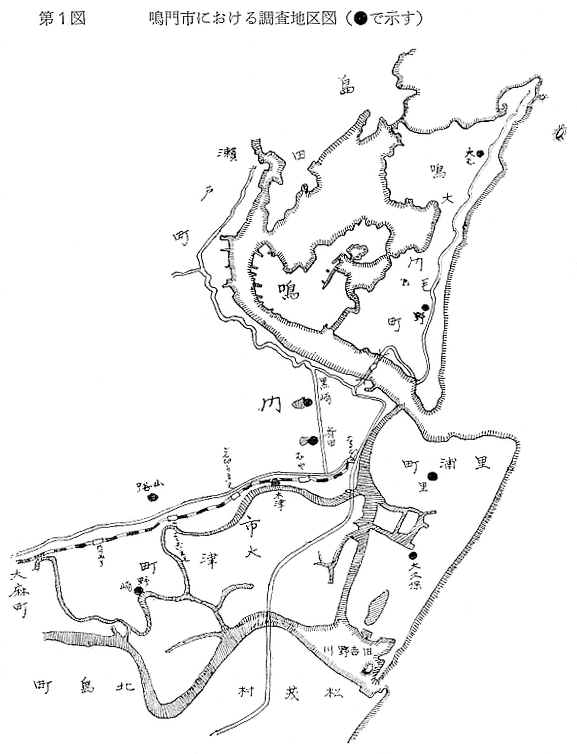
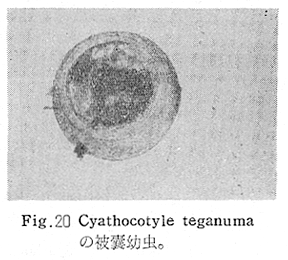
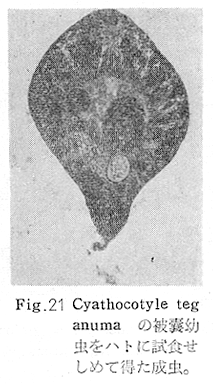
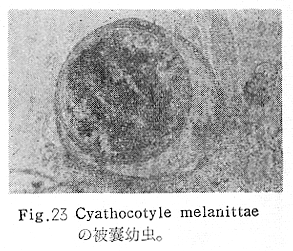
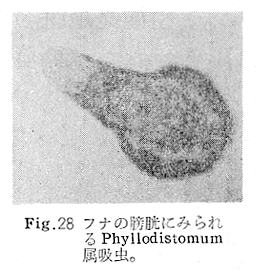
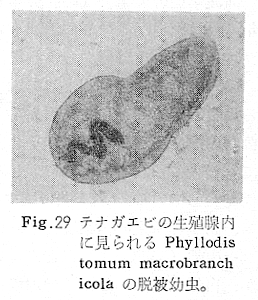
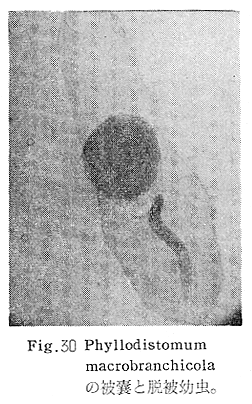
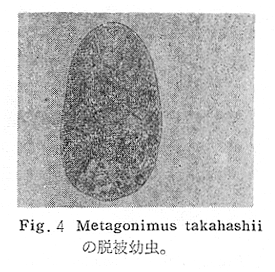
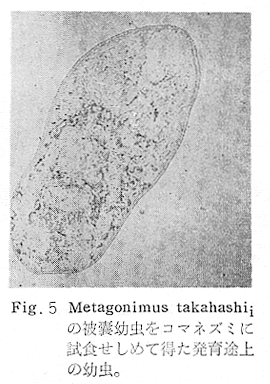
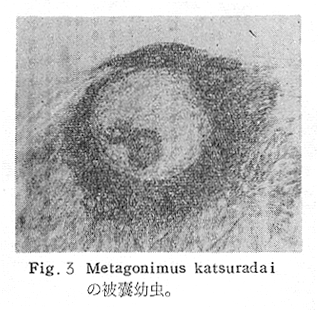
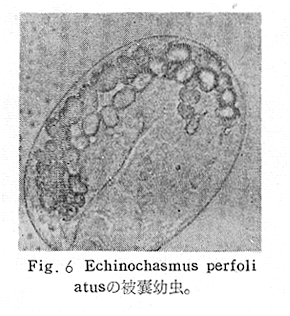
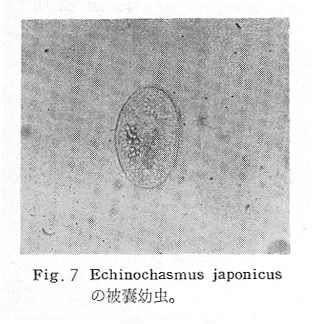
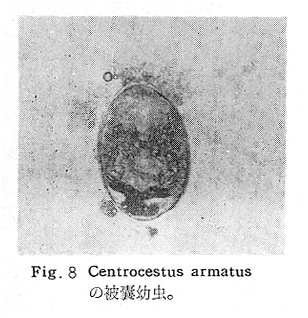
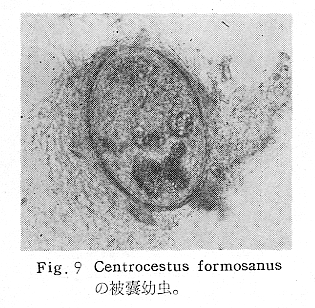
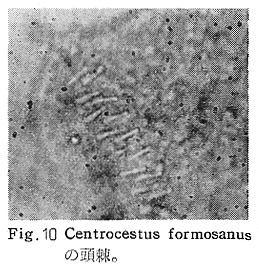
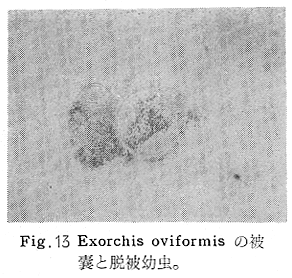
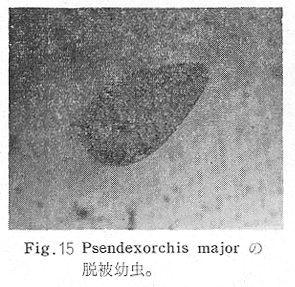
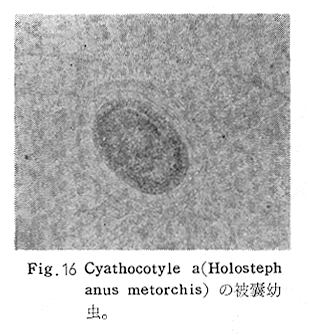
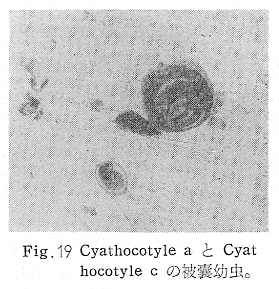
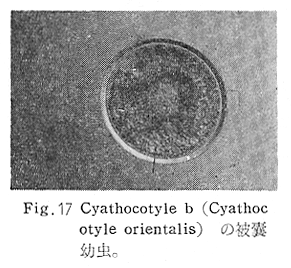
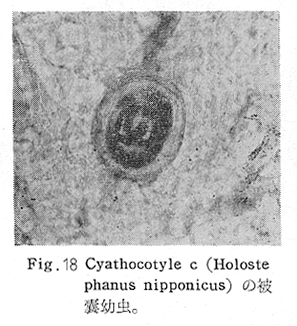
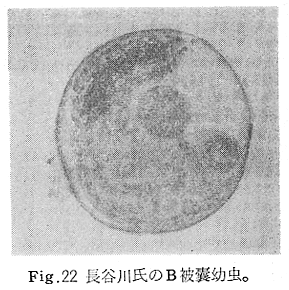
生物学班 豊岡磊造 一、はしがき 鳴門市の人体寄生虫については、村上邦平氏(1957)が大津町のモツゴを検査して、肝臓ジストマ、Metagonimus yokogawai,Echinochasmus perfoliatus,Echinochasmus japonicus,Centrocestus armatus の5種の人体寄生虫の被嚢幼虫を証明している。氏は人体寄生虫のほかに Exorchis oviformis,Pseudoexorchis major,Cyathocotyle a,Cyathocotyle b,Cyathocotyle c.長谷川氏の所属未定吸虫bの被嚢幼虫の寄生も認めている。同年、偶然にも筆者も同じ大津町のモツゴ、ハヤ(オイカワ)を検査して Metagonimus takahashii(Syn. Metagonimus yokogawai var. ovatus),Metagonimus katsuradai,Centrocestus armatus,Centrocestus formosanus,Echinochasmus perfoliatus,Echinochasmus japonicus の6種の人体寄生虫の被嚢幼虫を徳島生物学会2月例会(1957)に発表している。村上氏の得られた人体寄生虫の被嚢幼虫と筆者の得た被嚢幼虫を比較すると、次のようである。 (弟1表) 淡水魚を採集した場所は村上氏は大津町の徳長、木津、矢倉、備前島、大代、大幸の鉄道線路より南側の各地区にわたり詳細を極めているが、筆者のは鉄道線路の北側の山路地区のみである。しかし淡水魚の棲息する川は互に通じているので、大津町には上記8種類の人体寄生虫の被嚢幼虫が存在すると考えられる。筆者もその後、大津町のモツゴを検査して、Metagonimus yokogawai,Clonorchis sinensis の被嚢幼虫を認めることがきた。 鳴門旧市内については、村上氏はモツゴは全く棲息していないとして調査を打ち切っている。今回筆者は旧市内を中心として調査し、鳴門町、里浦町についても調査を試みた。調査の期間が少く、また炎熱下で、十分に調査することはできなかったが、今まで得た調査の結果を述べることとする。 二、調査地区と調査方法 調査を行った地区は鳴門旧市内の斎田、黒崎、鳴門町の大毛、野、里浦町の里、大久保、大津町の木津、山路、野崎の9地区である。採集は筆者が自分で行ない、網で捕獲した。捕獲した淡水魚はモツゴが主であるが、モツゴの棲息を認め得なかった地区ではフナ、ボラ等を捕獲した。 検査方法はウロコ、エラ、皮下組織および筋肉にわけ、スライドグラスで圧平し、双眼実体顕微鏡の下で精細にこれらを調査し、被嚢幼虫が認められると、これを顕微鏡下にうつし、倍率を拡大して種の検定を行った。生殖系原基などの観察および計測にはアセトカーミンで固定染色した材料を用いた。 三、調査成績 A、旧鳴門市内 旧鳴門市内の海岸地帯は塩田となっていて、淡水魚は認めることができなかったが、市街地と西部山地との間には相当広大な田園地帯があり、その地帯に給水するために、山際に大きな溜め池がところどころに存在し、そこから田園地帯に小さな溝が縦横に走っている。この溝の中にはフナ、ドジヨウなどの淡水魚が見られる。また溜め池にはモツゴ、フナがおびただしく棲息している。今回は主としてこれら溜め池のモツゴに重点を置いて調査した。捕獲したモツゴの数は1地区につき10数匹である。 (1)斎田地区 鳴門高校裏の溜め池(通称大池)のモツゴを調査したところ、5匹につき Echinochasmus japonicus は全個体に寄生が認められ、Cyathocotyle a はその中の4匹に寄生が認められた。E. japonicus が認められたことは、この池にマメタニシが棲息している証拠であり、肝臓ジストマの被嚢幼虫は認め得なかったが、軽度であっても肝臓ジストマの存在が予想される。河野圭典氏はこの池からマメタニシを採集している。 (第1図 鳴門市における調査地区図) (2)黒崎地区 黒崎小学校の後方、山際に大きな溜め池があり、この池の環境は前記斎田地区の大池と同じようである。この池のモツゴを採集して調査した結果、Echinochasmus japonicus,Centrocestus armatus,Cyathocotyle a,Cyathocotyle b の4種の被嚢幼虫が認められた。したがって、この池にもマメタニシおよび Centrocestus armatus の第1中間宿主であるカワニナが棲息していることは明らかであり、肝臓ジストマその他の人体寄生虫の感染源となり得る環境が十分に整えられている。 以上の調査結果よりして、旧市内の斎田、黒崎の山際には、人体寄生虫のマメタニシ、カワニナの第1中間宿主と、第2中間宿主のモツゴの棲息が認められたことよりして、旧鳴門市内もやはり肝臓ジストマその他の人体寄生虫の浸淫地としてこゝに付加したい。 B、里浦町 里浦通は北方は小高い山地になっていて海に面し、東方もまた海岸線となっている。旧吉野川の三角州としてできたものと考えられ、西北は旧鳴門市と堺し、西方には川を隔てて大津町がある。また南方は旧吉野川であり、その向うに松茂村がある。土質は砂地であり、甘藷の名産地である。甘藷畑には縦横に溝が走っており、ところどころに水門が設けられている。 (1)里地区 本地区にはモツゴの棲息は認められなかった。ボラ、フナは相当多数採集することができた。これらを精細に調査したが被嚢幼虫は全然見られなかった。 これは附近一帯が半鹹水であり、第1中間宿主のカワニナ、マメタニシ等の貝類の棲息不能のためと思われる。 (2)大久保地区 本地区は里地区の2粁ばかり南方に位置する。こゝでもモツゴは認められなかったが、ボラ、フナは相当多数棲息している。ボラには被嚢幼虫、成虫の吸虫寄生は認められなかったが、フナの肪胱には Phyllodistomum 属吸虫が認められた。これは筆者が海南町調査の折に、海部川下流地帯の塩原および浜崎産のフナから発見したものと同一であり、阿南市富岡町のフナにも見られる。またフナの筋肉には Exorchis oviformis の被嚢幼虫が見られた。この第1中間宿主はミズゴマツボであるが、この貝はカワニナ等の貝類よりも塩分濃度の高いところにおいても棲息できるもののようである。 C、鳴門町 鳴門町の調査は大毛島のみで島田島にまで足をのばすことはできなかった。大毛島にはモツゴは全くみられず、フナが少数見られたのみである。調査地区として、大毛、野をえらび、何れもフナを10数匹捕獲して詳細に剖検したが被嚢幼虫は全然認められなかった。これは第1中間宿主の貝類の棲息していないためであろう。 D、大津町 大津町は旧鳴門町の西方に位置し、北方は山地になっていて、南方は低く板野郡松茂村と境している。西方は大麻町と境されている。町内には大小無数の小川が流れており、淡水魚も豊富である。この地区は村上氏の詳細な報告があり、筆者も前に調査したところである。 (1)山路地区 本地区は大代の北方に位置し、その西北方には中山池があり、そこから田に水を引くために小川が本地区を横切り、大代地区に流れている。この小川の山際は梨畑であり、梨の消毒に使用した器具を洗条するために、小川の貝類、魚類は消毒時には殆んど死滅してしまう。しかし暫くすると、また下方から淡水魚がさかのぼってくる。消毒は主として初夏の候に行なわれるので、調査期と消毒期の期間が短かかったためか、今回の調査ではザリガニ、フナを認めただけである。前回の調査では「はしがき」で述べた如く、筆者はこゝのモツゴ、ハヤを検査して、Metagonimus takahashii,Metagonimus katsuradai,Echinochasmus perfoliatus,Echinochasmus japonicus,Centrocestus armatus,Centrocestus formosanus の6種の被嚢幼虫と長谷川氏のb被嚢幼虫を得ることができた。 (2)木津地区 木津地区は低温地帯であり、村上氏も述べられた如く、モツゴ、フナが非常に多い。この西方は旧鳴門市の南浜に接しているが、その境界附近で南浜側に「石越し」と名付けられる地点があり、その附近にはフナ、モツゴがおびただしく棲息している。しかし塩分濃度の大なるために第1中間宿主の貝類が少いためか、あるいは筆者の接集した個体が少いためか、「石越し」で採集したモツゴからは被嚢幼虫は認められなかった。ただ、フナには Metagonimus yokogawai の被嚢幼虫の寄生が認められた。少し上流の木津地区のモツゴからは肝臓ジストマ、Metagonimus yokogawai,Pseudoexorchis major,Cyathocotylee の4種の被嚢幼虫を得ることができた。 (3)野崎地区 本地区は大代の南方に位置し、低湿地帯であり、蓮根が沢山植えられている。旧吉野川に通ずる小川のモツゴ、ドンコ、テナガエビを検査したところ、モツゴから Metagonimus yokogawai,Exorchis oviformis,Ceotrocestus formosanus の3種、ドンコから種名不詳の1種、テナガエビからは Phyllodistomum macrobranchicola の被嚢幼虫をそれぞれ認めることができた。 四、従来の研究 四、五の項目については郷土研究発表会紀要および海南町誌には部分的にすでに述べたが、ここでは総括して述べることにする。 A)Metagonimus 属の吸虫ならびに被嚢幼虫 Metagonimus 属吸虫については、最初横川定氏(1911)が台湾において、アユより Metagonimus yokogawai の被嚢幼虫を発見し、これを犬に試食せしめて成虫となることを証明した。次で武藤昌知氏(1922)の研究により、その第1中間宿主がカワニナであることが証明された。後に高橋昌造氏(1929)はセルカリアについて詳しく研究し、主にアユに寄生しているのが Metagonimus yokogawai となるのであって、好んでフナ、金魚に寄生しているものはこれと異なり、卵も大きく、恐らく別種であろうといい Metagonimus takahashii Suzuki であると発表した。浅田順一氏(1934)は Metagonimus yokogawai と Metagonimus takahashii を比較研究し、後者を前者の変種であるとし、Metagonimus yokogawai var. ovatus(Kobayashi)とすべきことを論じた。勝田功夫氏(1932)は台湾においてボラに新被嚢幼虫を発見し、感染実験の結果、Metagonimus minutus Katsuta を得た。次に泉松之助氏(1935)は兵庫県下の淡水産魚類の被嚢幼虫を研究して、Metagonimus yokogawai に似ているが排出嚢が小型で、下正Y字型をした Metagonimus 属被嚢幼虫を発見し、これを哺乳類に試食させて成虫とし、Metagonimus katsuradai Izumi と命名した。Metagonimus yokogawai と Metagonimus takahashii の区別については、後者を独立の1種とすべきか、あるいは前者の変種とすべきか、色々議論があり、その決定は今後に残されている。被嚢幼虫についても、宿主によって区別できる場合とできない場合がある(岡部浩洋氏、1940)というが、筆者はこれをここでは種として取り扱い、Metagonimus takahashii として、浅田氏の述べた次の特徴によって、両者を分類した。 Metagonimus yokogawai……体は著しく透明にして淡黄色、弱拡大にて膀胱著明に黒色を呈す。 Metagonimus takahashii……体は前者に比して透光性に乏しく、色は帯黄褐色、弱拡大にて膀胱の黒色著明ならず。 B)Centrocestus 属の吸虫ならびに被嚢幼虫 Centrocestus 属吸虫の被嚢幼虫は最初田部浩氏(1922)がオイカワ、カワムツなどから発見し、これを犬、ネコに与えて成虫とし、Stamnosoma armatum Tanabe と名付けた。氏は魚類を好んで生食するゴイサギ、アオサギおよびネコにおいて自然寄生を認めている。更に氏は実験的に自らこの被嚢幼虫を試食し、人体にも感染可能であることを立証した。翌年(1923)錦織正雄氏は台湾において、Stamnosoma armatum とは別種の Stamnosoma formosanum を発見し、その発育史についても研究して、第1中間宿主はカワニナであり、第2中間宿主はコイ科、ナマズ科、ドジヨウ科など13種の淡水魚であり、終宿主は実験的には犬、ネコ、ウサギ、シロネズミなど、ほとんどすべての実験動物に感染可能であると述べている。その後、黒川帝文氏(1929)により、Stamnosoma armatum の第1中間宿主もやはりカワニナであることが明らかにされた。黒川氏はまた広島県下住民中より Stamnosoma 属吸虫を発見し、これを Stamnosoma formosanum と比較して、宿主および頭棘数の相異からその変種であるとして、Stamnosoma farmosanum var. kurokawai と命名した(1934)。次で泉松之助氏(1935)は兵庫県下の淡水魚5種から Stamnosoma 属被嚢幼虫を発見し、これを Stamnosoma nycticoracis Izumi と名付けている。その後 Stamnosoma 属は山口左仲氏等により Centrocestus 属に編入されて、Stamnosoma armatum は Centrocestus armatus,Stamnosoma formosanum は Centrocestus formosanus という様に属名が変更された。Centrocestus formosanus,Centrocestus formosanus var. kurokawai,Centrocestus nycticoracis の主な相異点は頭棘数と宿主であるが、頭棘数は計算違いと思われる点があり、また宿主も実験方法によれば寄生不可能とされた動物でも寄生可能である点(郷土研究発表会紀要6・7・8合併号)よりして、これは本来同一種であるようであるので、ここでは最初に命名した錦織氏の Centrocestus formosanus を採ることにした。 C)Echinochsmus 属の吸虫ならびに被嚢幼虫 Rutz(1908)はハンガリーにおいて、犬、猫の小腸より本属吸虫を発見して、Echinochasmus perfoliatus と命名した。本邦において最初にこれを記載したのは田部浩氏(1915)であり、氏は淡水魚を犬、ネコ、人に試食させて感染することを証明した。第1中間宿主は武藤昌知氏(1917)により、マメタニシであることが確かめられた。田部浩氏(1926)は Echinochasmus perfoliatus に似て、大きさの極めて小さい本属吸虫を発見し、これを Echinochasmus japonicus と名付けた。氏によるとこの被嚢幼虫はタナゴ、オイカワ、ドンコ等19種の淡水魚に見られ、猫に感染させて成虫とした。氏家直記氏(1936)は本被嚢幼虫を嚥下して人体にも感染可能なることを証明した。この第1中間宿主はマメタニシであるが、長谷川恒治氏(1929)によると、「Echinochasmus perfoliatus の卵は被嚢幼虫を感染せしめて第18日目に糞便中にあらわれるのに反して、Echinochasmus japonicus の卵は第9日目にあらわれ、武藤氏のマメタニシのセルカリアをモツゴに感染させて得た被嚢幼虫を犬、ネコに試食せしめると、第8日目に成虫が得られるので、武藤氏が Echinochasmus perfoliatus のセルカリアとして発表されたものは実は Echinochasmus japonicus のものであろう。」と述べている。 D)Exorchis 属および Pseudoerorchis 属の吸虫ならびに被嚢幼虫 小林晴次郎氏(1915)はナマズの膀胱から本属吸虫を発見し、これを Exorchis oviformis と名付けた。岡部浩洋氏(1936)はミズゴマツボ Stenothyra japonica(Hirase M. S.Kuroda)よりこのセルカリアを発見し、これを金魚に試食させて本属被嚢幼虫となることを証明した。また翌年オタマジヤクシが本吸虫の第2中間宿主になり得ることを発表した。 Pseudoexorchis major については、最初長谷川恒治氏(1927)がアユの鱗中に本被嚢幼虫を発見し、ナマズを終宿主とすることを予報した。高橋昌造氏(1929)はカワニナに寄生するセルカリアを魚類に感染させて、この被嚢幼虫となることを証明した。後に長谷川氏(1935)はこれに Exorchis major なる種名を与えた。山口左仲氏(1938)はモツゴに寄生する本種被嚢幼虫をナマズに試食させて成虫を得るとともに、これに Pseudoexorchis なる属名を与えて独立させた。 E)Cyathocotyle 科の吸虫ならびに被嚢幼虫 Cyathocotyle 科の被嚢幼虫については長谷川恒治氏(1934)が岡山県においてモツゴを検査してa、b、cの3種を発見している。山口左仲氏(1939、1940)は長谷川氏のa、b、cの型に属する Cyathocotyle 科の被嚢幼虫の発育史をそれぞれ追求して、a種は Holostephanus metorchis、b種は Cyathocotyle orientalis、c種は Holostephanus nipponicus に発育させている。また氏はこれら3種とは異なった Cyathocotyle 科の1新被嚢幼虫をモツゴより見出し、これをトビに試食させて成虫とし、Cyathocotyle melanittae に発育させている(1941)。そしてその成虫の自然寄生をビロウドキンクロの小腸内に認めている(1934)。筆者は(1962)Cyathocotyle melanittae の被嚢幼虫として山口氏が記載したもの(第20図)をハトを用いて感染実験を行ったところ、全く別種の Cyathocotyle teganuma(第21図)に発育した。この成虫は石井信太郎氏(1929)が千葉県手賀沼産のカイツブリの腸から発見したものである。また筆者は徳島市においてフナの筋肉内に1新被嚢幼虫(第23図)を認め、これを同じくハトを用いて感染実験を行ったところ、Cyathocotyle melanittae(第24図)に発育した(1961)。 F)Phyllodistomum 属の吸虫ならびに被嚢幼虫 本属吸虫については、山口左仲氏(1953)によれば38種あり、本邦産のものとしては5種を挙げている。このうち淡水産および汽水産のものは次の3種である。 Phyllodistomum parasilusi yamaguti,1934……ナマズに寄生、京都。 P. mogurundae yamaguti,1934……ドンコに寄生、京都。 P. macrobranchicola yamaguti,1934……メタセルカリアはテナガエビに寄生、成虫はドンコに感染実験をして得られる。自然宿主は不明。 筆者は(1962)海部川下流地帯の塩原および浜崎産フナの膀胱内より Phyllodistomum 属の吸虫を発見した。これはまだ成虫に達していなかったが、その後阿南市富岡町産フナを調査したところ、完全に発育した成虫が認められた。今回の調査では里浦町大久保産フナに同様の未成熟の吸虫(第28図)を見出すことができた。また大津町野崎産テナガエビを検査したところ、その生殖腺内に Phyllodistomum macrobranchicola の被嚢幼虫(第29、30図)が認められた。 Phyllodistomum 属吸虫のセルカリアについては、長船寛治氏(1898)が岡山においてマシジミより1種の長尾セルカリアを発見している。山口左仲氏(1953)はビワ湖産セタシジミより、長船氏の記載したものと同じと思われるセルカリアを見出し、それが多分 Phyllodistomum の1種に発育するものであろうと述べている。筆者は吉野川および小松島産マシジミおよびヤマトシジミを調査して、Phyllodistomum 属セルカリアを2種見出すことができた(1954)。その1種は上記長船氏、山口氏の記載したものと同じと思われるが、他の1種は尾が短く、明らかに別種である。またこのセルカリアはスポロシスト中においてメタセルカリアに発育しているものが見られた。これらについては理科学会誌に発表の予定である。 五、各種被嚢幼虫の鑑別 被嚢幼虫の特徴については、多くの著者によりたびたび述べられている。ここに述べるのは筆者が実際に徳島県下において採集した魚類に寄生する被嚢幼虫について観察したもので、他にも宿主となり得る魚類が存在するはずである。また被嚢幼虫の種類についても徳島県下には、ここに述べる以上の種類が認められるが、ここでは筆者が鳴門市において今までに認め得たものに限定して記述することにする。 (1)Clonorchis sinensis の被嚢幼虫 モツゴの筋肉、皮下組織、鱗片に寄生する。 被嚢はだ円形を呈し、大きさは縦径0.15ミリメートル、横径0.12ミリメートルである。被嚢の壁は2層よりなり、透明であり、外層は弾力性をもち強靱であるが、内層はうすく弱々しい。嚢内においては幼虫は体を2重に折りまげて存在する。幼虫と被嚢壁の間には空隙が存在し、そこには粘液様物質が認められる。 脱皮幼虫は体表に微棘をもち、殊に体前半部において著しい。口吸盤は体の前端腹面に開口し、縦径0.043、横径0.042ミリメートルある。腹吸盤は口吸盤より大きく、縦径0.048ミリメートル、横径0.046ミリメートルある。口吸盤につづき短い前咽頭がある。咽頭は縦径0.025、横径0.024ミリメートルある。食道は細長く、体の中央附近で二岐して腸管となる。腸管は体の後端附近で盲管に終っている。消化管内には円板状の球状体を含んでいる。排出嚢は腹吸盤以下の体後部の大部分を占め、嚢状で中に黒色顆粒を含んでいる。 (2)Metagonimus yokogawai の被嚢幼虫(第1図) オイカワ、モツゴ、ヤリタナゴ、カワムツの鱗片、ひれ、皮下組織にみられる。 被嚢は鱗片におけるものは円形、ひれにおけるものはだ円形である。被嚢の大きさは鱗片におけるもので直径約0.15ミリメートルである。嚢内における幼虫は普通体をまげており、盛に旋回運動をしている。幼虫の表面には小さな棘をもっている。体色は淡黄色を呈し、口吸盤、咽頭、食道、腹吸盤が認められる。排出嚢は後体部に位置し黒色顆粒が著明である。 (3)Metagonimus takahashii の被嚢幼虫(第2、4、5図) フナ、ドジヨウ、オイカワ、モツゴ、ヤリタナゴ、カワムツの鱗片、ひれ、皮下組織に寄生する。 本被嚢幼虫の被嚢は M.yokogawai のと同様、鱗片におけるものは円形で、直径0.15ミリメートルぐらいである。ひれにおけるものは長だ円形である。被嚢の壁は2層よりなり、ガラス様透明である。被嚢内の幼虫は体を2重に折りまげて存在する。 脱皮幼虫は体表に小さな棘を有し、大きさは体の伸縮の状態により異るが、縦径0.29ミリメートル、横径0.12ミリメートルぐらいである。体の先端に口吸盤があり、大きさは縦径0.03ミリメートル、横径0.05ミリメートルある。腹吸盤は体中央附近の左側に存在し、大きさは縦径0.025、横径0.015ミリメートルある。口吸盤に続いて短い前咽頭があり、咽頭となっている。咽頭は縦径0.02ミリメートル、横径0.025ミリメートルである。これに続いて食道があり、間もなく分岐して左右の両腸脚となっている。腸脚は体両側を後方に走り、体の後端附近で盲管に終っている。排出嚢は体後端部を占め、中に裸粒を含んでいる。 (4)Metagonimus katsuradai の被嚢幼虫(第3図) オイカワ、モツゴ、ヤリタナゴの鱗片、ひれに寄生する。 被嚢は円形またはだ円形であり、大きさは直径0.18ミリメートルぐらいである。被嚢壁は無色透明で、内外2属よりなる。被嚢中の虫体は体を二重に折って存在し、時々わずかに運動する。 脱皮幼虫は縦径0.20ミリメートル、横径0.12ミリメートルぐらいあり、体表には微棘をもっている。先端には口吸盤があり、大きさは縦径0.035ミリメートル、横径0.03ミリメートルである。腹吸盤は体中央右側に偏して存在し、直径0.021ミリメートルである。口吸盤に続いて非常に短い前咽頭があり、咽頭となる。咽頭は大きさ縦径0.02ミリメートル、横径0.015ミリメートルである。咽頭に続いて長い食道があり、分岐して腸管となり、体後端附近で盲管に終っている。消化管の内部には円盤状の球状体が多数存在し、消化管内を移動している。睾丸の原基は体後端近くで排出嚢を隔てて相対している。卵巣原基は体の正中線上で排出嚢の直前に位置する。排出嚢は前二者と異なりY字型である。 (5)Echinochasmus perfoliatus の被嚢幼虫(第6図) オイカワ、モツゴの鰓葉に寄生する。 被嚢はだ円形を呈し、大きさは縦径0.07〜0.086ミリメートル、短径0.054〜0.05ミリメートルである。被嚢の壁は2層よりなり、ガラス様透明である。嚢中における幼虫は体軸を長軸に一致させるか、または体を2つに折って存在し、わずかに運動している。 脱被幼虫は体表に微棘を有する。体前端部には口吸盤があり、大きさは縦径0.04ミリメートル、横径0.03ミリメートルあり、腹吸盤は体後端より体長の3分の1の所に位置し、大きさは縦径0.04ミリメートル、横径0.05ミリメートルあり口吸盤より少し大きい。口吸盤の周囲には一列をなして24本の頭棘がある。口吸盤につづいて短い前咽頭があり咽頭となる。咽頭は縦径0.025ミリメートル、横径0.02ミリメートルある。食道は細長く腹吸盤の前端附近で左右の両腸脚に分岐する。排出嚢は特有の形をしていて、体後部の嚢状部から2本の排出管が前方に伸び、腹吸盤の附近で幅が広くなり、咽頭の両側附近で細管となっている。そして中に50個位の顆粒を含んでいる。排出嚢には赤色の色素がみられる。 (6)Echinochasmus japonicus 被嚢幼虫(第7図) オイカワ、モツゴの鰓葉に寄生する。 被嚢は前者に類似するが、小形であり、大きさは縦径0.075〜0.078ミリメートル、横径0.046〜0.057ミリメートルである。被嚢の壁は内外2層よりなり、弾力性が強い。幼虫は被嚢内では体軸を被嚢の長軸に一致させて存在する。 脱皮幼虫は体表に微棘を有し、虫体には赤色色素が認められない。口吸盤は前端にあり、縦径0.02、横径0.025ミリメートルある。腹吸盤は体後端より前方3分の1の所に位置し、口吸盤と同じ大きさである。口吸盤の附近には24本の頭棘が1列にならんでいる。消化管の構造は E. perfoliatus に類似し、咽頭の大きさは直径0.015ミリメートルある。排出嚢の形も前種に似るが、顆粒は20個あまりを有するに過ぎない。 (7)Centrocestus armatus の被嚢幼虫(第8図) オイカワ、モツゴ、カワムツの肝臓に多いが、内臓壁や筋肉、鰓葉にも寄生する。 被嚢はだ円形であり、大きさは縦径0.17〜0.19ミリメートル、横径0.11ミリメートルある。被嚢の壁は透明な内外2層からなり、両端が肥厚している。幼虫は被嚢の長軸と一致して体を動かす。 脱皮幼虫は大きさ縦径0.35ミリメートル、横径0.20ミリメートルである。口吸盤は前端にあり、周囲に2列になって合計44本の頭棘を有する。前咽頭は極めて短く、咽頭の大きさは縦径0.03ミリメートル、横径0.025ミリメートルである。これに続いて食道があり、間もなく分岐して腸脚となる。腸脚は排出嚢の前方において盲管に終っている。腸の内部には円板状の球状体が見られる。腹吸盤は体中央部附近に位置し、直径0.04ミリメートルある。排出嚢は工字型を呈し、黒色の顆粒を含んでいる。排出嚢の両側には睾丸の原基が認められる。 (8)Centrocestus・formosanus の被嚢幼虫(9、10、11、12図) フナ、ドジョウ、オイカワ、モツゴ、ヤリタナゴ、タモロコ、メダカの鰓葉基部、軟骨組織間の間隙に寄生する。 被嚢はだ円形を呈し、大きさは縦径0.24ミリメートル、横径0.16ミリメートルある。被嚢の壁は2層よりなり、両端は肥厚していない。嚢内における幼虫は被嚢の長軸に一致して、やや体を短縮して存在する。 脱皮幼虫は体表に微棘を有し、縦径0.28ミリメートル、横径0.15ミリメートルある。口吸盤は体前端にあり、直径0.05ミリメートルある。口吸盤の周囲には2列に頭棘が取り巻き、合計42本ある。腹吸盤は体中央部附近に位置し、口吸盤より小型であり、0.035ミリメートルある。前咽頭は短く、咽頭は縦径0.036、横径0.018ミリメートルある。食道は間もなく分岐して腸脚となり、排出嚢の前端にて盲管に終っている。腸管内には円板状の球状体がみられる。排出嚢は不正I字型を呈し、その両側に睾丸の原基が認められる。 (9)Erorchis oviformis 被嚢幼虫(第13、14図) フナ、モツゴの筋肉、鰓、ひれ、皮下組織に見られる。 被嚢はだ円形を呈し、大きさは縦径0.2ミリメートル、横径0.16ミリメートルある。 脱皮幼虫はダルマ型を呈し、体表は微棘で覆われている。口吸盤は先端近くに位置し、大きさは直径0.045ミリメートルある。短い前咽頭についで、だ円形の咽頭があり、大きさは縦径0.028ミリメートル、横径0.023ミリメートルである。食道は非常に短く、直ちに分岐して腸管となる。腸管は体の後端附近で盲管に終っている。腹吸盤は小さく体の中央部より少しく前端に位置し、大きさは直径0.03ミリメートルある。咽頭の少し下方両側に眼点を有する。眼点は一側に1個、他の側に2個存在する。腹吸盤の少し下方で腸脚の両側には大きな睾丸がある。腹吸盤に接して生殖系原基らしい細胞塊が見られるが、恐らく貯精嚢に発育するものであろう。咽頭と腹吸盤の間には小さな腺細胞が沢山存在する。排出嚢は大きく、V字型またはY字型であり、その先端は咽頭附近にまで達する。中に黒色顆粒を入れている。 (10)Pseudoexorchis mayor の被嚢幼虫(第15図) モツゴ、ヤリタナゴ、カワムツ、タモロコの鱗、ヒレ、鰓に寄生する。 被嚢の大きさは縦径0.22ミリメートル、横径0.17ミリメートルある。被嚢の壁は内外2層からなっているが、うすく破れ易い。嚢内の幼虫は普通体を2つに折って存在する。 脱皮幼虫は縦径0.3ミリメートル,横径0.18ミリメートルあり、体表は微棘で覆われている。口吸盤は大きく、縦径0.065〜0.069ミリメートル、横径0.078〜0.082ミリメートルあり、すぐ咽頭となる。咽頭は大きさ直径0.021ミリメートルある。咽頭についで短い食道があり腹吸盤の前方において左右の腸脚にわかれる。腸は睾丸にまで達する。腹吸盤は小さく、直径0.039ミリメートルである。睾丸は大きく、体後端部の両側に相対して左右相称的に位置する。右睾丸の上部には卵巣原基がみられる。排出嚢はY字型である。食道の両側には眼点がある。眼点附近から体後端附近にかけて黄褐色の色素が散在している。 (11)Cyathocotylea(Holostephanus metorchis)の被嚢幼虫(第16、19図) モツゴの筋肉に寄生する。 被嚢は宿主から由来したと考れられる繊維性の袋で包まれている。この袋の内層は透明で結合組織様のものからなり、一層でできている。被嚢と虫体の間には可なりのスペースがあり、たくさんの小さな球形の顆粒で満されている。 脱皮幼虫は縦径0.25ミリメートル、横径0.2ミリメートルあり、体表には微棘を有する。口吸盤は先端にあり、直径0.06ミリメートルある。咽頭は縦径0.03ミリメートル、横径0.032ミリメートルである。腹吸盤は縦径0.025ミリメートル、横径0.03ミリメートルであり、その後方に大きな吸着器がみられる。排出嚢は半円形の2つの輪が正中線の両側にあり、それが咽頭の背面でたがいにくっついている。 (12)Cyathocotyle b(Cyathocotyle orientalis)の被嚢幼虫(第17図) モツゴ、フナの筋肉に寄生する。 被嚢は円形を呈し、大きさは直径0.25ミリメートルである。被嚢の壁は2層からなり、外側の層は宿主から由来したと考えられる厚さ0.01ミリメートルぐらいの結合組織の層からできており、内側の層は厚い透明なガラス様物質からなっている。虫体は被嚢内に充満して存在し、口吸盤、咽頭、吸着器が認められる。体の大部分は排出嚢で占められている。 脱皮幼虫は縦径0.2〜0.25ミリメートル、横径0.15〜0.20ミリメートルあり、前端には口吸盤があり、大きさは直径0.04ミリメートルある。腸吸盤は縦径0.018ミリメートル、横径0.02ミリメートルあり、吸着器の前方に存在する。吸着器は直径0.08〜0.09ミリメートルある。排出嚢は腹側から観察すれば単純な輪にみえるが、側方から観察すれば2重の輪であることがわかる。 (13)Cyathocotyle c(Holostephanus nipponicus)の被嚢幼虫(第18、19図) モツゴの筋肉内に寄生する。 被嚢は宿主から由来したと考えられるだ円形の厚い結合組織の袋で包まれている。被嚢と袋との間にはかなり厚い黄褐色の顆粒が存在する。被嚢は円形を呈し、直径0.35ミリメートルあり、Cyathocotyle a にくらべると大きい。被嚢の壁は同じ厚さの2層よりなっている。被嚢と虫体の間には虫体が排出した黒色の顆粒状の排出物で満されている。 脱被幼虫は縦径0.23ミリメートル、横径0.17ミリメートルあり、体表に微棘を有する。口吸盤は直径0.05ミリメートル、咽頭は縦径0.025ミリメートル、横径0.028ミリメートルある。食道の部分は認められず、ただちに分岐して腸管となり、体後端附近で盲管に終っている。腹吸盤は直径0.02ミリメートルあり、吸着器の前方に存在する。吸着器は大きく、直径0.08ミリメートルある。排出嚢の形は Cyathocotyle a の場合と殆んど同じであるが、2つの輪の結合点の両側から前方に小さな突起がでている。 (14)長谷川氏のB被嚢幼虫(第22図) モツゴ、オイカワの肝臓および腎臓に寄生する。 球形を呈した大きな被嚢幼虫であり、被嚢の大きさは直径0.52ミリメートルある。被嚢の壁は比較的うすく、幅約0.013ミリメートルある。被嚢は2層からなっており、内層はうすくガラス様であり、外層は厚く繊維様である。この被嚢は宿主から由来したと考えられる結合組織に包まれている。嚢内における幼虫は弓状に体をまげて存在し、時々僅かに運動する。スライドとカバーの間で微圧をかければ、幼虫は容易に脱被する。 脱被幼虫は体に微棘を有する。口吸盤は球形で直径0.091ミリメートルあり、比較的短い前咽頭に続いて小さな咽頭がある。腸管は体の後端附近で盲管に終っている。腹吸盤は体中央よりも少しく後端に位置し、口吸盤よりも大きく直径0.126ミリメートルある。口吸盤の後方両側には数個の腺細胞があり、口吸盤の先端附近で外に開口している。排出嚢はI字型で腹吸盤の後端附近に達し、中に小さな多くの顆粒を入れている。体後端から腹吸盤の附近にかけて生殖細胞の原基がみられる。 (15)筆者の所属未定種(第25、26、27図) ドンコの筋肉内に寄生する。 被嚢はだ円形を呈し、大きさは縦径0.31ミリメートル、横径0.23ミリメートルある。被嚢の壁は透明なうすい層でできている。 脱被幼虫はだ円形を呈し、縦径0.386ミリメートル、横径0.286ミリメートルである。体前端部に口吸盤があり、大きさは縦径0.09ミリメートル、横径0.065ミリメートルある。口吸盤に続いて短い前咽頭がある。咽頭は縦径0.022ミリメートル、横径0.036ミリメートルあり、短い食道を経て分岐して腸脚となる。腸の長さは023ミリメートル、幅は0.018ミリメートルである。生殖系原基は2ケ所に分散して存在する。体後端の正中線附近に存在するものは大きく、0.054×0.068ミリメートルあり、体中央部の右側方附近に存在するものは小さく0.022×0.029ミリメートルである。腹吸盤は認められない。 Phyllodistomum 属の吸虫ならびに被嚢幼虫、Cyathocotyle 科の他の種類については稿を改めて述べることにする。 六、まとめ 鳴門市の旧市内、大津町、里浦町、鳴門町の淡水産魚類を調査した結果、次の結論を得た。 1、従来旧市内にはモツゴ、マメタニシの棲息が認められないとされていたが、旧市内の山際にはこの棲息が認められ、肝臓ジストマその他の人体寄生虫の感染地としての環境がととのえられている。 2、里浦町は半鹹水の地点が多く、人体寄生虫は認められない。Exorchis oviformis の被嚢幼虫と Phyllodistomum 属の成虫がフナに認められる。 3、鳴門町にはフナは棲息しているが、ジストマは認められない。 4、大津町には肝臓ジストマ、Metagonimus yokogawai 等8種の人体寄生虫の被嚢幼虫が認められる。 5、大津町産ドンコには一新被嚢幼虫が認められる。またテナガエビには Phyllodistomum macrobranchicola の被嚢幼虫が認められる。 主要文献 錦織正雄(1924)、すたむのぞーま属ノ一新種竝ニ其発育史ニ就テ、病理学誌14、526〜531、 田部 浩(1926)、淡水産魚類ヲ中間宿主トスル吸虫類の研究、第三、ゑきのかすむす属、 一新種 Echinochasmus japonicus n. sp.、日本医学会誌第16年:295〜299、 栗栖荘太郎(1931)、我国の鳥類を終末宿主とする吸虫類の研究、特に Corves coronoides japonensis Bonapate に於て認められたる吸虫の三種に就て、東医誌2740:1929〜1938。 ―(1931)、我国の沢蟹に於て新に発見したる一種の被嚢幼虫について、東医誌2735:1683〜1687。 石井信太郎(1932)、鳥類寄生吸虫類の研究(第一報)、実験医誌16:1205〜1321。 ―(1932)、鳥類寄生吸虫類の研究(第二報)、実験医誌19:467〜479。 yamaguti, S.(1934)、Studies on the helminth fauna of Japan. Part2,Trematodes of fishes, 1, Jap. Zool. 5(3) 泉 松之助(1935)、兵庫県下に於ける淡水産魚類を中間宿主とする吸虫類の研究、東医新誌2949:10〜11。 ―(1935)、「スタムノゾーマ」属吸虫の一種竝其の発育史に関する研究、東医新誌2948:17〜24。 yamaguti, S.(1935)、Studies on the helminth fauna of Japan. Part25,Trematodes of birds,6, Jap. J. Zool. 8(2)165〜167。 小林久雄(1935)、日本淡水産魚類竝に其寄生虫、養賢堂。 岡部浩洋(1936)、Exorchis ouiformis Kobayashi(Trematoda)ノ発育史ニ就テ、福岡医誌29(1):211〜220。 ―(1938)、The second intermediate hosts of Exorchis ouiformis kobayashi. Fukuoka Acta med. 30:106〜109。 ―(1939)、北満農業移民地における淡水魚を中間宿主とする吸虫類に就て、福岡医大誌:289〜296。 ―(1940)、福岡県下ニ於ケル淡水産魚類ヲ中間宿主トスル吸虫類ノ被嚢幼虫総覧、福岡医誌33(3)、309〜335 Yamaguti, S.(1942)、Studies on the helminth fauna of Japan. Part, 38. Larval trematodes of fishes, Jap. J. Med. Sc 6 Bacteriol. & Parasitol. Yamaguti, S.(1953)、Systema Helminthum, Part1, Digenetic trematodes of fish. Maruzen Co。 豊岡磊造(1954)、シジミに寄生するセルカリアに就て、日本動物学会中国四国支部会報No6:11。 山口富雄(1956)、四国においてはじめて見出したマメタニシについて、(肝吸虫1)、四国医誌9、4:220〜221。 村上邦平(1957)、徳島県下の肝吸虫の研究、とくに第2中間宿主の分布について、四国医誌10、1:11〜31。 豊岡磊造(1957)、徳島県における淡水産魚類の被嚢幼虫(第1報)、実験生物学報(学会記事)、7、3:127。 ―(1959)、徳島県における淡水産魚類の被嚢幼虫(第2報)、実験生物学報(学会記事)、9、6:251。 ―(1961)、徳島県における淡水産魚類の被嚢幼虫について、郷土研究発表会紀要(6、7、8合併号):15〜26。 ―(1962)、海部川下流地帯における淡水産魚類の吸虫ならびに被嚢幼虫、海南町誌:32〜38。 |
|
| 徳島県立図書館 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}